

Phage-assisted evolution of an adenine base editor with improved Cas domain compatibility and activity
- PMID: 32433547
- PMCID: PMC7357821
- DOI: 10.1038/s41587-020-0453-z
Phage-assisted evolution of an adenine base editor with improved Cas domain compatibility and activity
Erratum in
-
Author Correction: Phage-assisted evolution of an adenine base editor with improved Cas domain compatibility and activity.Nat Biotechnol. 2020 Jul;38(7):901. doi: 10.1038/s41587-020-0562-8. Nat Biotechnol. 2020. PMID: 32433548
Abstract
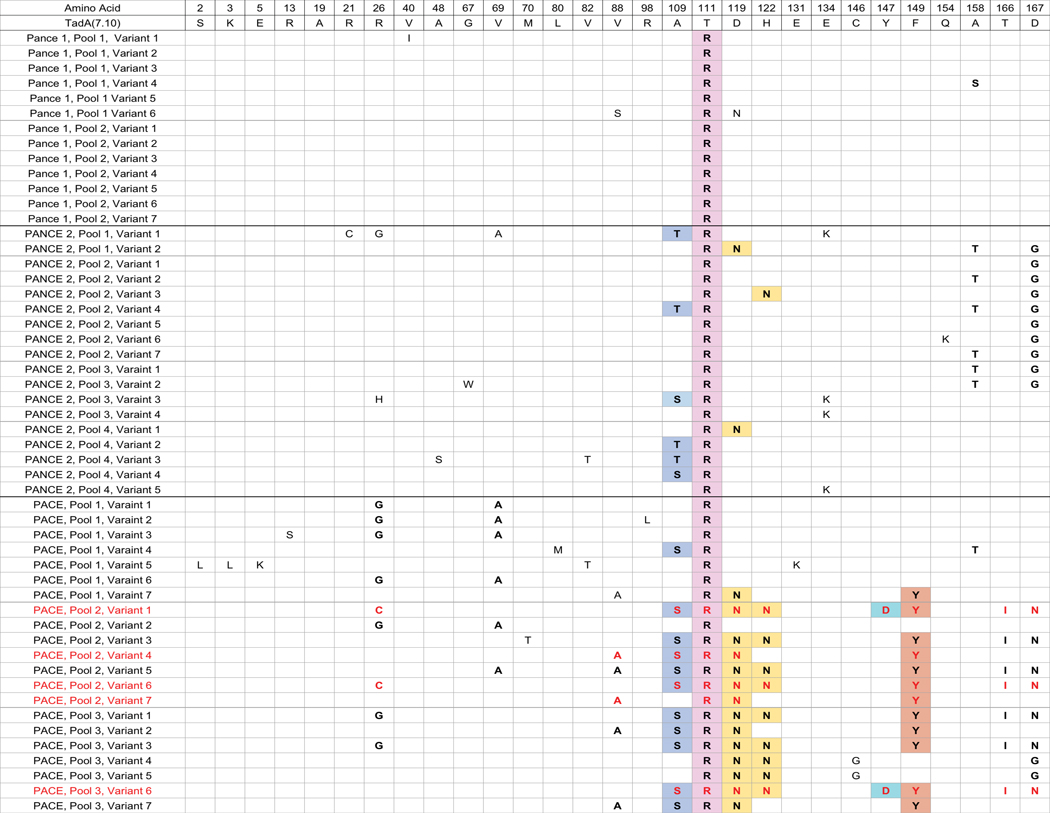
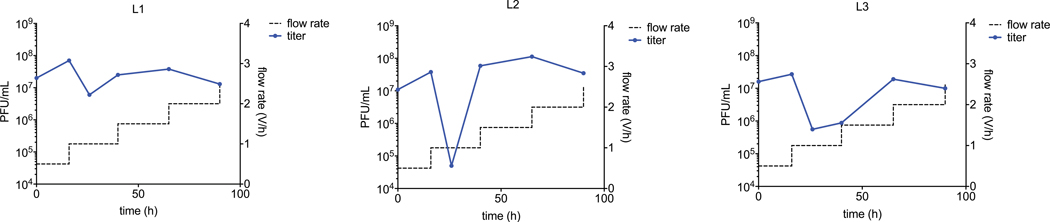
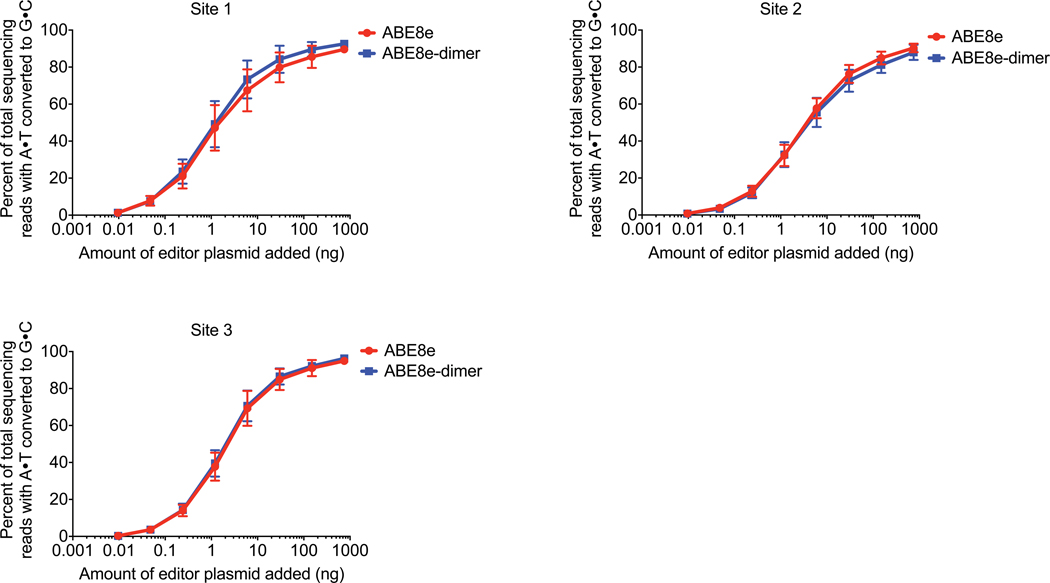
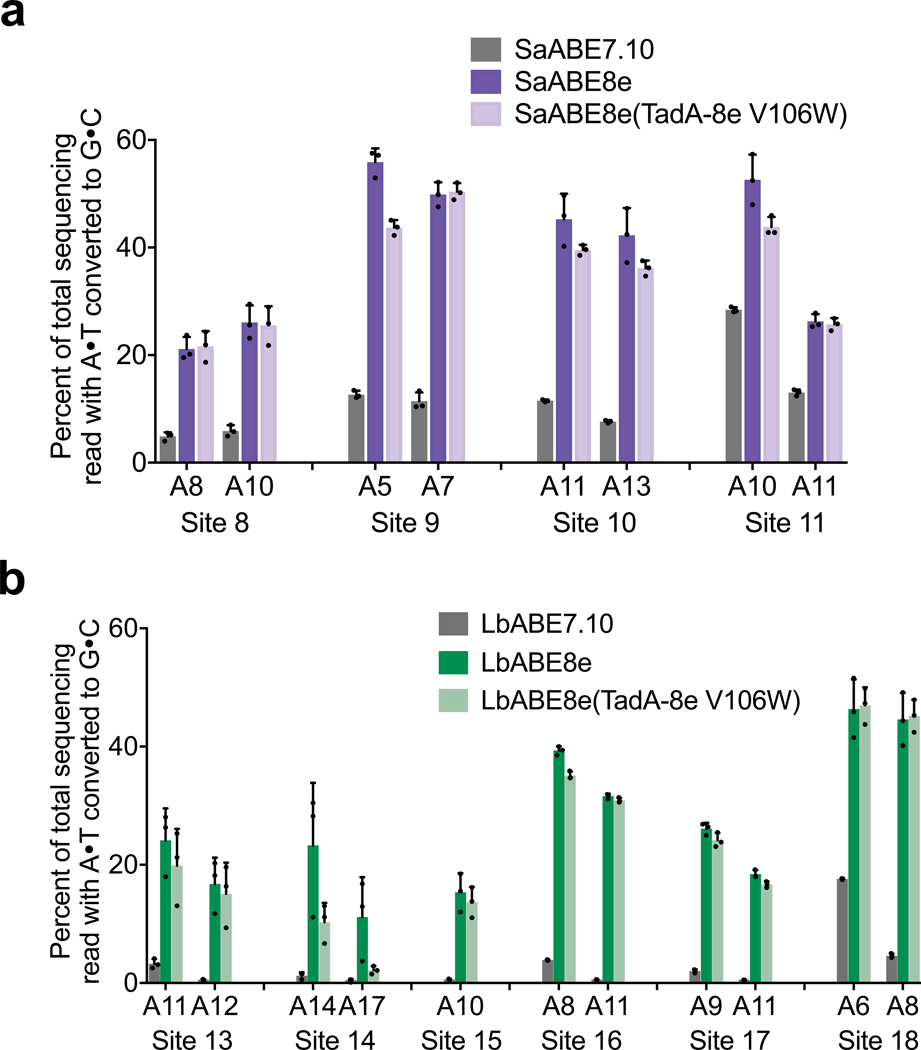
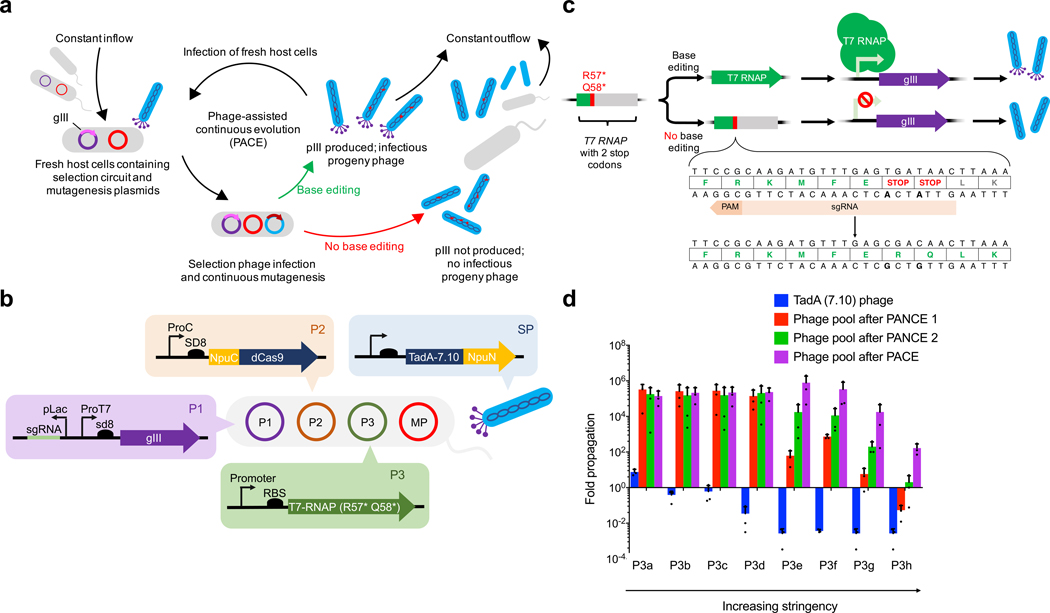
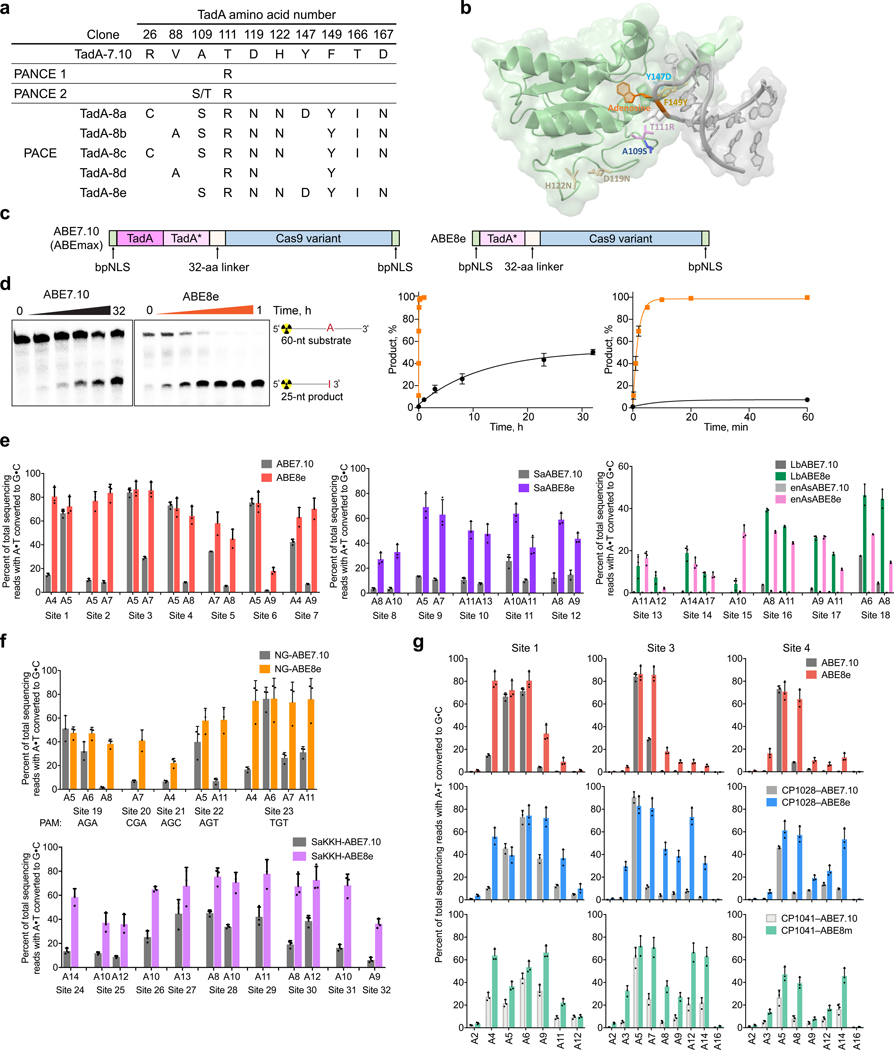
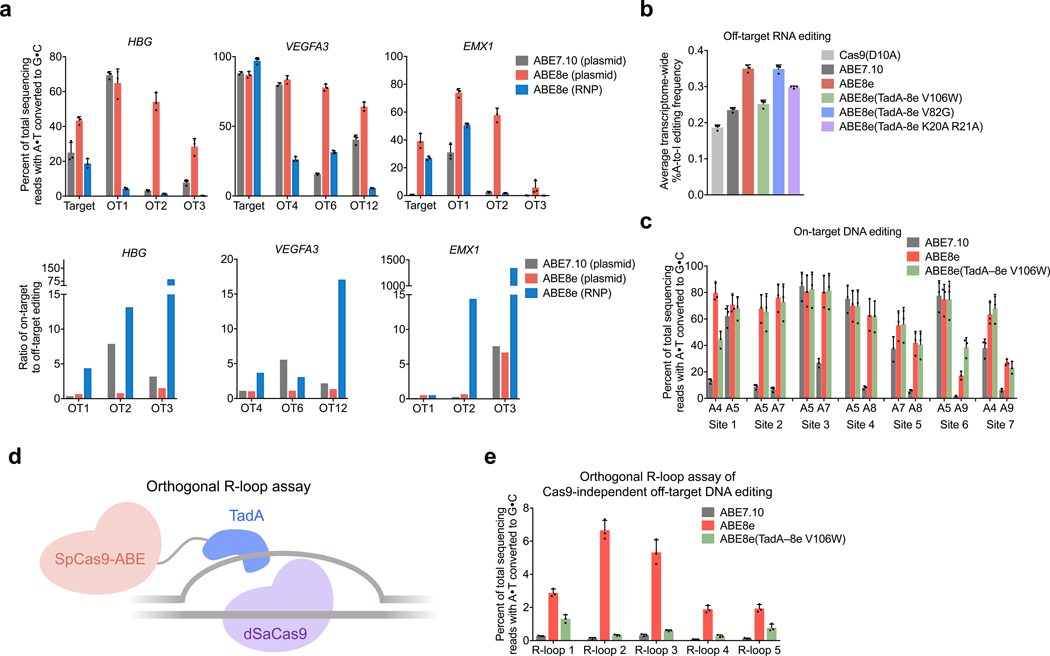
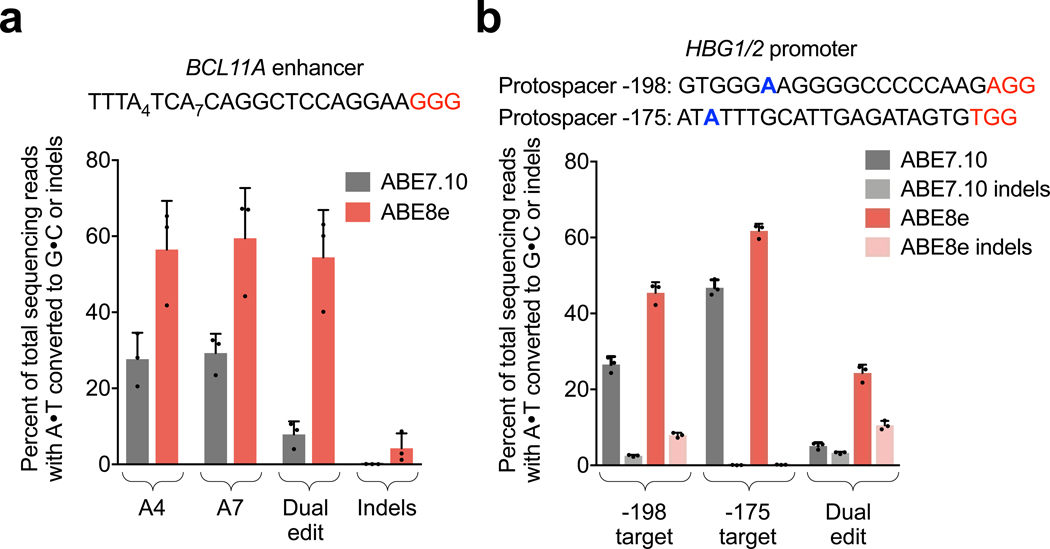
Applications of adenine base editors (ABEs) have been constrained by the limited compatibility of the deoxyadenosine deaminase component with Cas homologs other than SpCas9. We evolved the deaminase component of ABE7.10 using phage-assisted non-continuous and continuous evolution (PANCE and PACE), which resulted in ABE8e. ABE8e contains eight additional mutations that increase activity (kapp) 590-fold compared with that of ABE7.10. ABE8e offers substantially improved editing efficiencies when paired with a variety of Cas9 or Cas12 homologs. ABE8e is more processive than ABE7.10, which could benefit screening, disruption of regulatory regions and multiplex base editing applications. A modest increase in Cas9-dependent and -independent DNA off-target editing, and in transcriptome-wide RNA off-target editing can be ameliorated by the introduction of an additional mutation in the TadA-8e domain. Finally, we show that ABE8e can efficiently install natural mutations that upregulate fetal hemoglobin expression in the BCL11A enhancer or in the the HBG promoter in human cells, targets that were poorly edited with ABE7.10. ABE8e augments the effectiveness and applicability of adenine base editing.
Figures
References
-
- Nishida K. et al. Targeted nucleotide editing using hybrid prokaryotic and vertebrate adaptive immune systems. Science 353 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
