

Discordant rearrangement of primary and anamnestic CD8+ T cell responses to influenza A viral epitopes upon exposure to bacterial superantigens: Implications for prophylactic vaccination, heterosubtypic immunity and superinfections
- PMID: 32433711
- PMCID: PMC7239382
- DOI: 10.1371/journal.ppat.1008393
Discordant rearrangement of primary and anamnestic CD8+ T cell responses to influenza A viral epitopes upon exposure to bacterial superantigens: Implications for prophylactic vaccination, heterosubtypic immunity and superinfections
Abstract
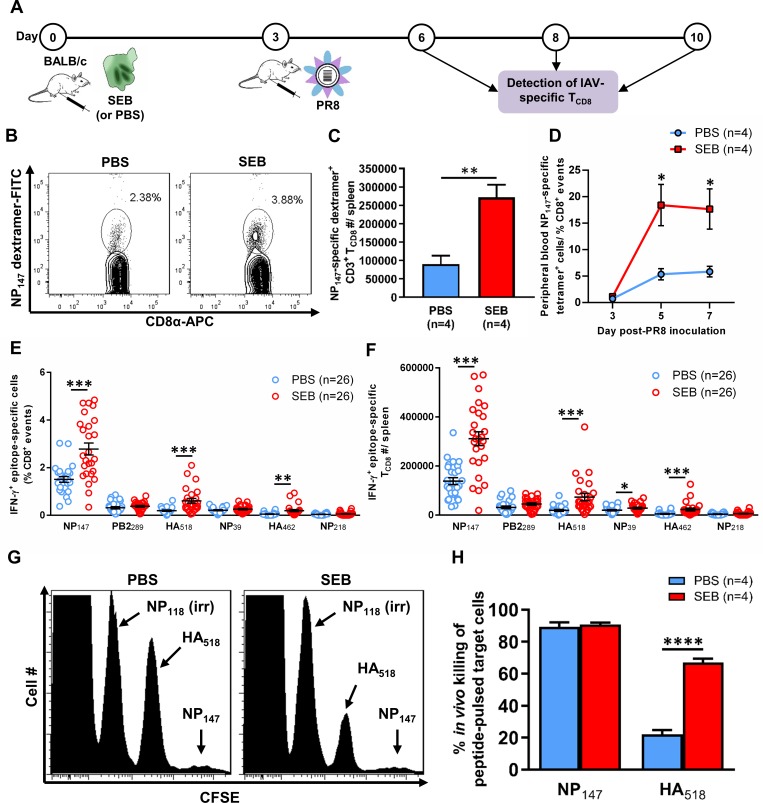
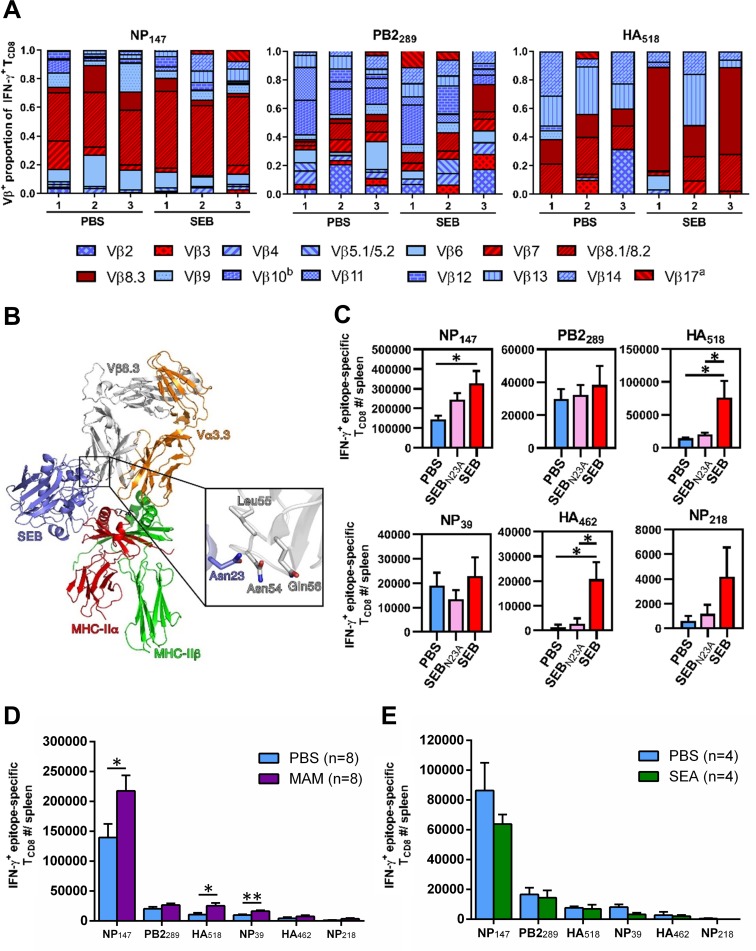
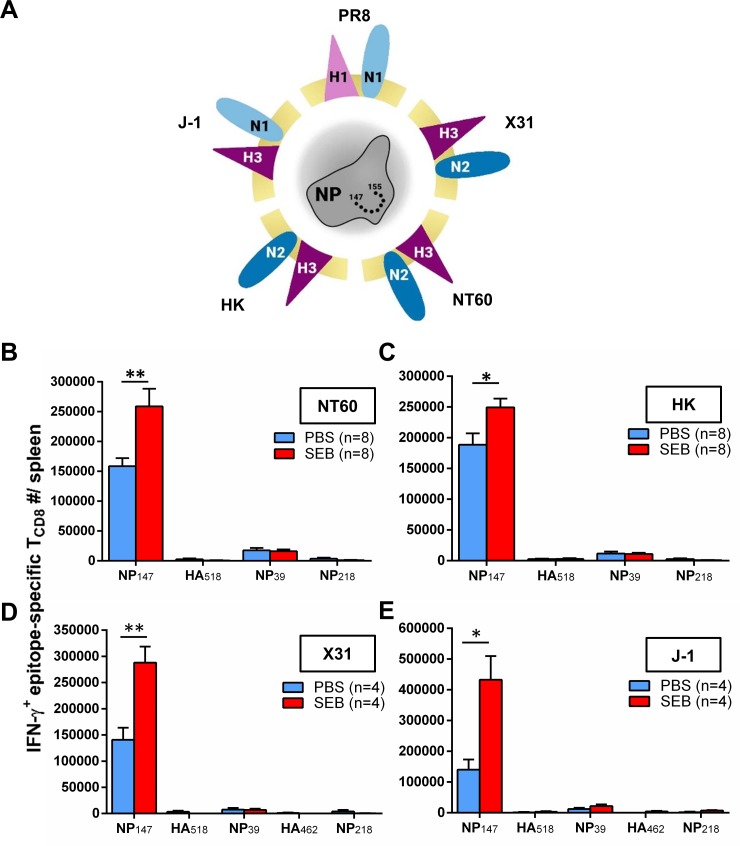
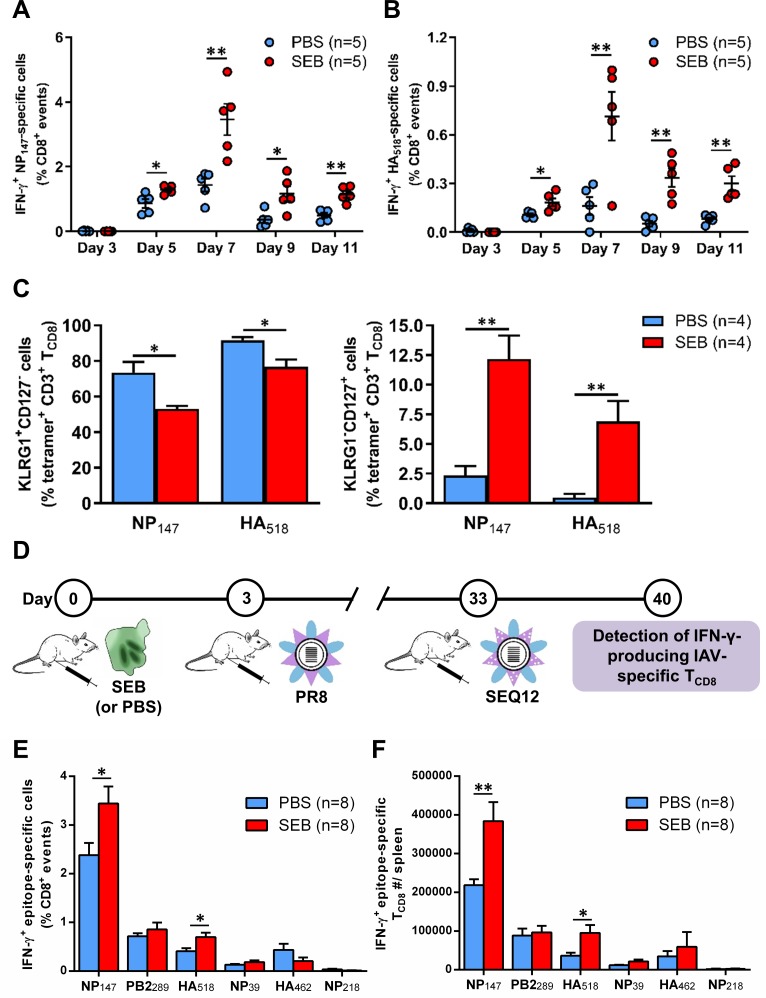
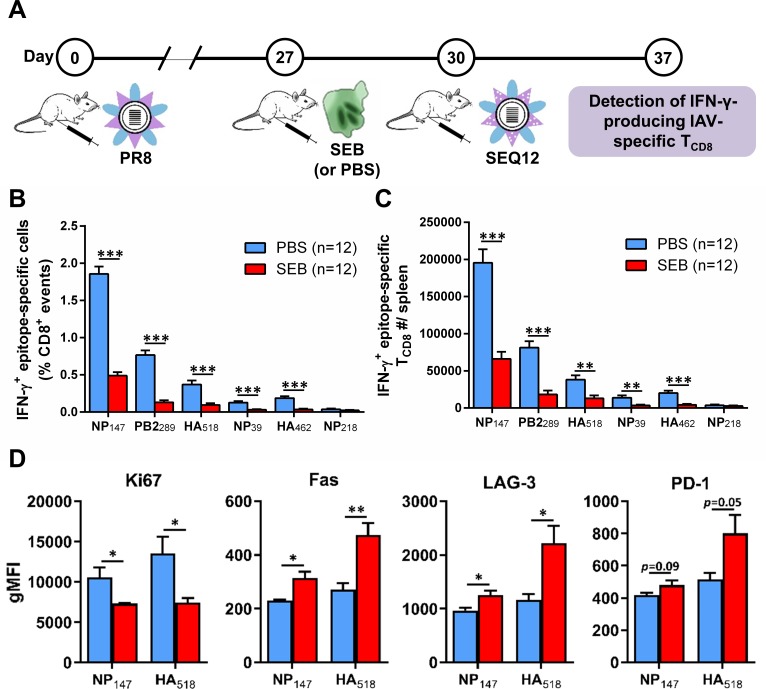
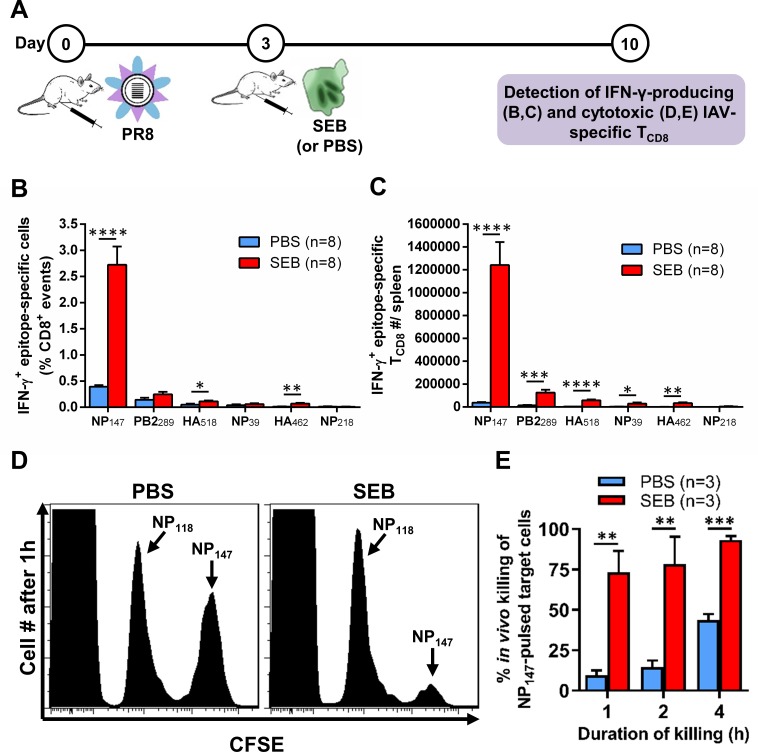
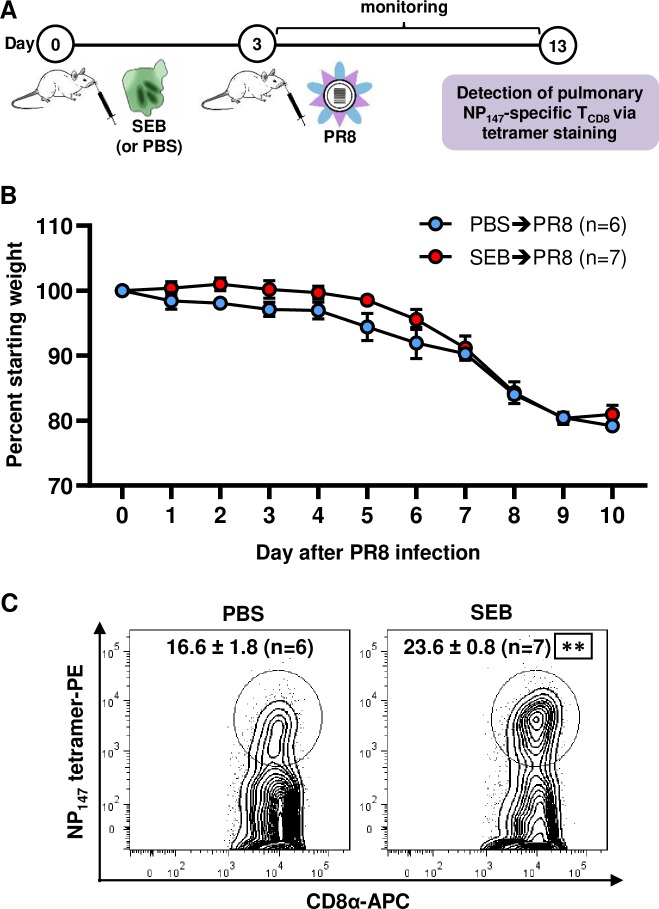
Infection with (SAg)-producing bacteria may precede or follow infection with or vaccination against influenza A viruses (IAVs). However, how SAgs alter the breadth of IAV-specific CD8+ T cell (TCD8) responses is unknown. Moreover, whether recall responses mediating heterosubtypic immunity to IAVs are manipulated by SAgs remains unexplored. We employed wild-type (WT) and mutant bacterial SAgs, SAg-sufficient/deficient Staphylococcus aureus strains, and WT, mouse-adapted and reassortant IAV strains in multiple in vivo settings to address the above questions. Contrary to the popular view that SAgs delete or anergize T cells, systemic administration of staphylococcal enterotoxin B (SEB) or Mycoplasma arthritidis mitogen before intraperitoneal IAV immunization enlarged the clonal size of 'select' IAV-specific TCD8 and reshuffled the hierarchical pattern of primary TCD8 responses. This was mechanistically linked to the TCR Vβ makeup of the impacted clones rather than their immunodominance status. Importantly, SAg-expanded TCD8 retained their IFN-γ production and cognate cytolytic capacities. The enhancing effect of SEB on immunodominant TCD8 was also evident in primary responses to vaccination with heat-inactivated and live attenuated IAV strains administered intramuscularly and intranasally, respectively. Interestingly, in prime-boost immunization settings, the outcome of SEB administration depended strictly upon the time point at which this SAg was introduced. Accordingly, SEB injection before priming raised CD127highKLRG1low memory precursor frequencies and augmented the anamnestic responses of SEB-binding TCD8. By comparison, introducing SEB before boosting diminished recall responses to IAV-derived epitopes drastically and indiscriminately. This was accompanied by lower Ki67 and higher Fas, LAG-3 and PD-1 levels consistent with a pro-apoptotic and/or exhausted phenotype. Therefore, SAgs can have contrasting impacts on anti-IAV immunity depending on the naïve/memory status and the TCR composition of exposed TCD8. Finally, local administration of SEB or infection with SEB-producing S. aureus enhanced pulmonary TCD8 responses to IAV. Our findings have clear implications for superinfections and prophylactic vaccination.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Similar articles
-
Bacterial Superantigens Expand and Activate, Rather than Delete or Incapacitate, Preexisting Antigen-Specific Memory CD8+ T Cells.J Infect Dis. 2019 Apr 8;219(8):1307-1317. doi: 10.1093/infdis/jiy647. J Infect Dis. 2019. PMID: 30418594 Free PMC article.
-
MAIT cells launch a rapid, robust and distinct hyperinflammatory response to bacterial superantigens and quickly acquire an anergic phenotype that impedes their cognate antimicrobial function: Defining a novel mechanism of superantigen-induced immunopathology and immunosuppression.PLoS Biol. 2017 Jun 20;15(6):e2001930. doi: 10.1371/journal.pbio.2001930. eCollection 2017 Jun. PLoS Biol. 2017. PMID: 28632753 Free PMC article.
-
Severity of Acute Infectious Mononucleosis Correlates with Cross-Reactive Influenza CD8 T-Cell Receptor Repertoires.mBio. 2017 Dec 5;8(6):e01841-17. doi: 10.1128/mBio.01841-17. mBio. 2017. PMID: 29208744 Free PMC article.
-
Human influenza viruses and CD8(+) T cell responses.Curr Opin Virol. 2016 Feb;16:132-142. doi: 10.1016/j.coviro.2016.01.016. Epub 2016 Mar 12. Curr Opin Virol. 2016. PMID: 26974887 Review.
-
[Viral superantigens].Mikrobiyol Bul. 2016 Jul;50(3):491-504. doi: 10.5578/mb.24250. Mikrobiyol Bul. 2016. PMID: 27525406 Review. Turkish.
Cited by
-
Targeting the MR1-MAIT cell axis improves vaccine efficacy and affords protection against viral pathogens.PLoS Pathog. 2023 Jun 29;19(6):e1011485. doi: 10.1371/journal.ppat.1011485. eCollection 2023 Jun. PLoS Pathog. 2023. PMID: 37384813 Free PMC article.
-
Novel insights into the immune response to bacterial T cell superantigens.Nat Rev Immunol. 2024 Jun;24(6):417-434. doi: 10.1038/s41577-023-00979-2. Epub 2024 Jan 15. Nat Rev Immunol. 2024. PMID: 38225276 Review.
-
T Cell Immunity to Bacterial Pathogens: Mechanisms of Immune Control and Bacterial Evasion.Int J Mol Sci. 2020 Aug 26;21(17):6144. doi: 10.3390/ijms21176144. Int J Mol Sci. 2020. PMID: 32858901 Free PMC article. Review.
-
In Vivo Cytotoxicity by α-GalCer-transactivated NK Cells.Methods Mol Biol. 2021;2388:157-174. doi: 10.1007/978-1-0716-1775-5_15. Methods Mol Biol. 2021. PMID: 34524671
-
The superantigens SpeC and TSST-1 specifically activate TRBV12-3/12-4+ memory T cells.Commun Biol. 2023 Jan 20;6(1):78. doi: 10.1038/s42003-023-04420-1. Commun Biol. 2023. PMID: 36670205 Free PMC article.
References
-
- Peters T, Hammon D, Jarrah R, Palavecino E, Blakeney E et al. Staphylococcal Toxic Shock Syndrome Complicating Influenza A Infection in a Young Child. ISRN Pulmonology. 2011;1–3.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
