

New GMP manufacturing processes to obtain thermostable HIV-1 gp41 virosomes under solid forms for various mucosal vaccination routes
- PMID: 32435515
- PMCID: PMC7235025
- DOI: 10.1038/s41541-020-0190-9
New GMP manufacturing processes to obtain thermostable HIV-1 gp41 virosomes under solid forms for various mucosal vaccination routes
Abstract
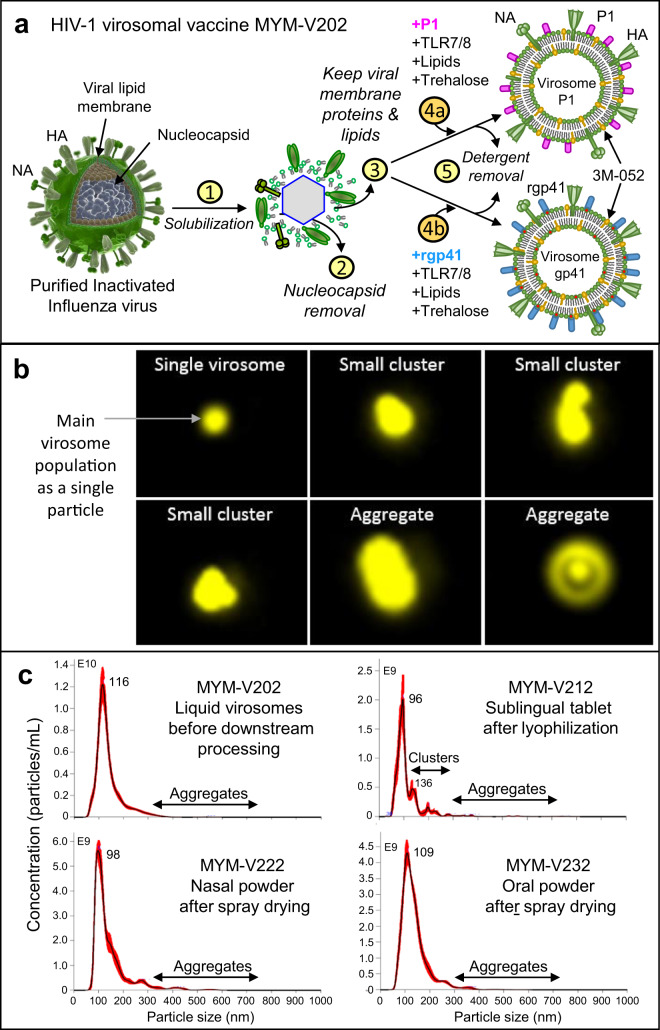
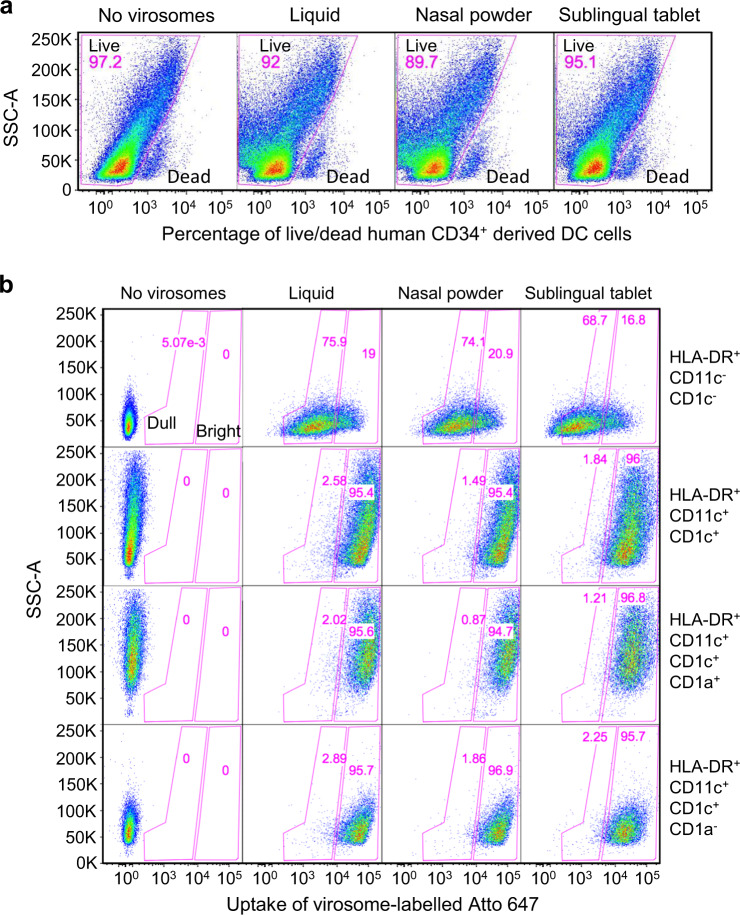
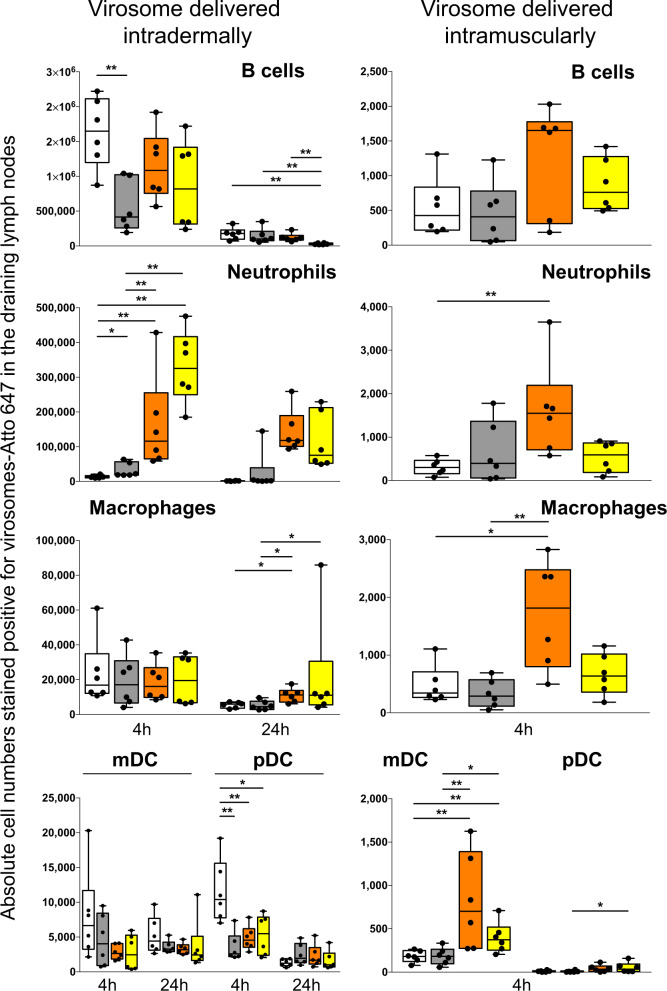
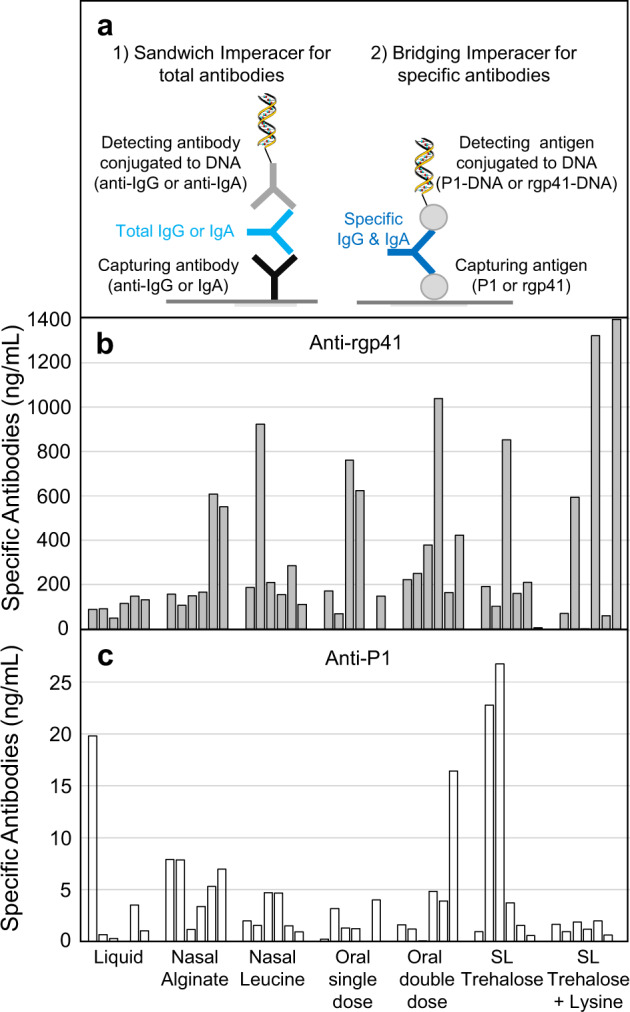
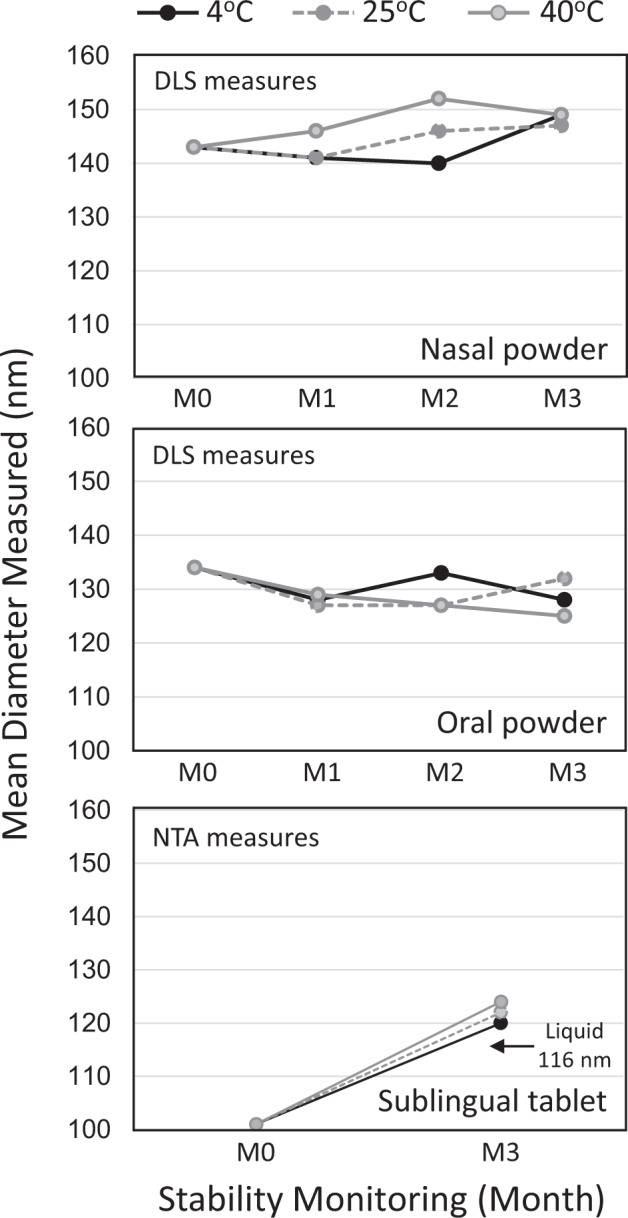
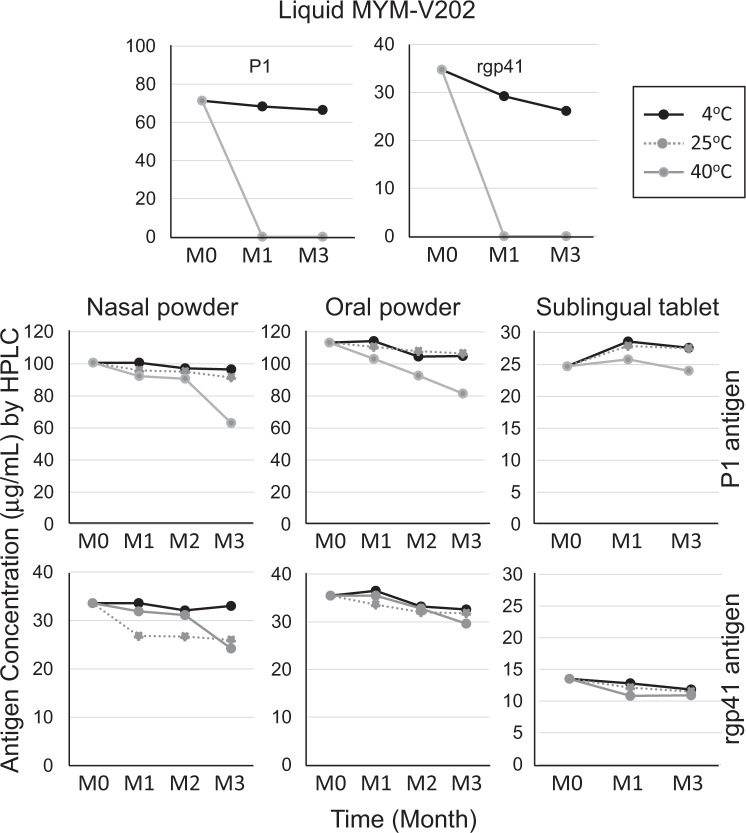
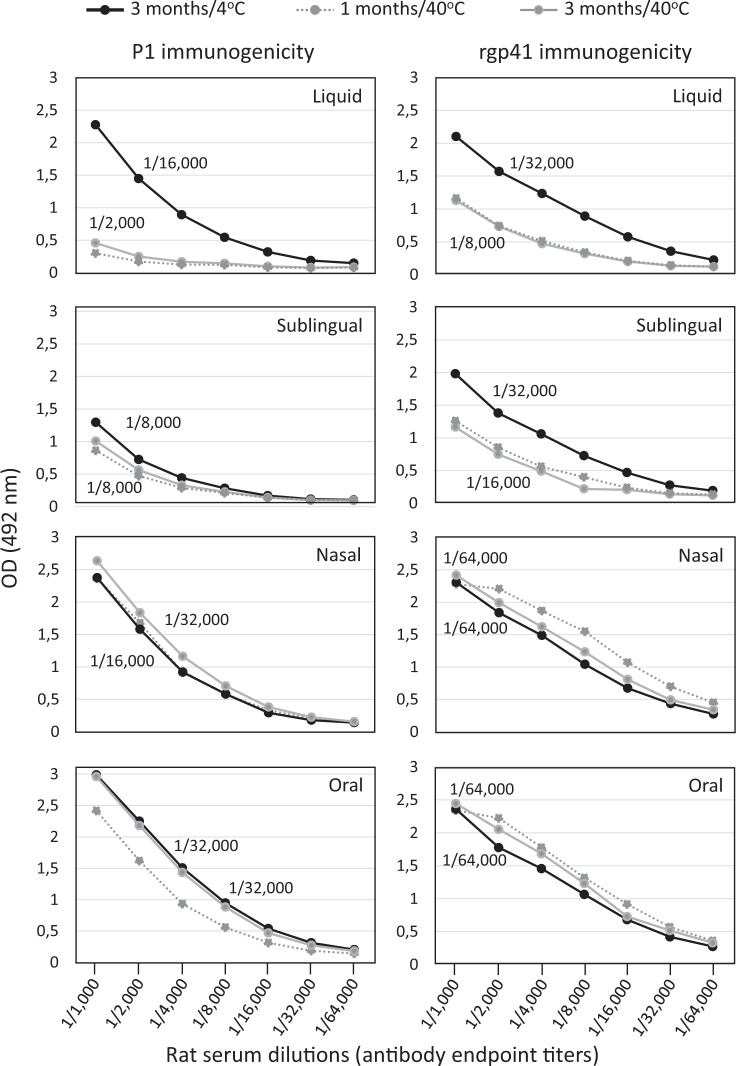
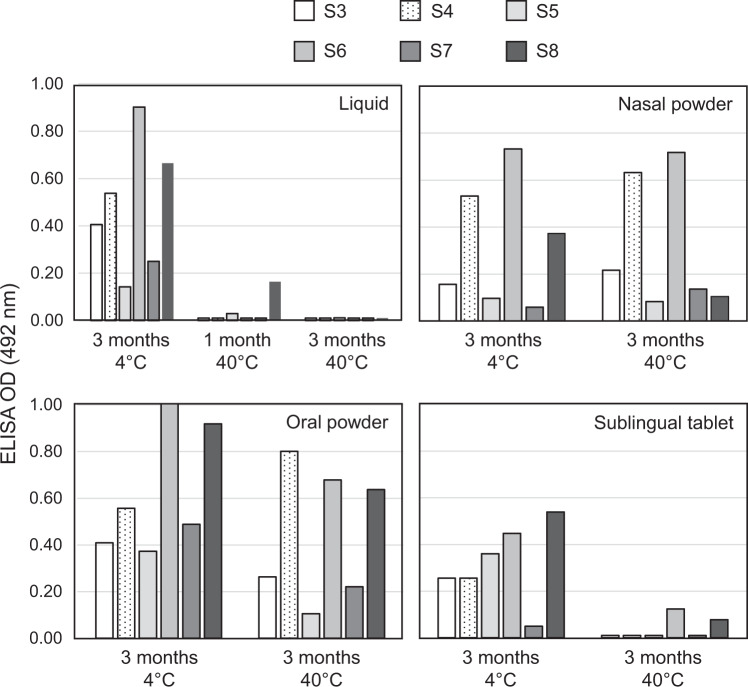
The main objective of the MACIVIVA European consortium was to develop new Good Manufacturing Practice pilot lines for manufacturing thermostable vaccines with stabilized antigens on influenza virosomes as enveloped virus-like particles. The HIV-1 gp41-derived antigens anchored in the virosome membrane, along with the adjuvant 3M-052 (TLR7/8 agonist) on the same particle, served as a candidate vaccine for the proof of concept for establishing manufacturing processes, which can be directly applied or adapted to other virosomal vaccines or lipid-based particles. Heat spray-dried powders suitable for nasal or oral delivery, and freeze-dried sublingual tablets were successfully developed as solid dosage forms for mucosal vaccination. The antigenic properties of vaccinal antigens with key gp41 epitopes were maintained, preserving the original immunogenicity of the starting liquid form, and also when solid forms were exposed to high temperature (40 °C) for up to 3 months, with minimal antigen and adjuvant content variation. Virosomes reconstituted from the powder forms remained as free particles with similar size, virosome uptake by antigen-presenting cells in vitro was comparable to virosomes from the liquid form, and the presence of excipients specific to each solid form did not prevent virosome transport to the draining lymph nodes of immunized mice. Virosome integrity was also preserved during exposure to <-15 °C, mimicking accidental freezing conditions. These "ready to use and all-in-one" thermostable needle-free virosomal HIV-1 mucosal vaccines offer the advantage of simplified logistics with a lower dependence on the cold chain during shipments and distribution.
Keywords: Biotechnology; Immunology; Infectious diseases.
© The Author(s) 2020.
Conflict of interest statement
Competing interestsM. Amacker and S.F. are employees of Mymetics SA, T.S. and F.B. are employees of Mymetics BV. S.F. and T.S. own equity in the mother company Mymetics Corporation. Mymetics Corporation shareholders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. R.J. is an employee of Upperton Ltd. and owns equity in Upperton Ltd. Mymetics Corporation and Catalent U.K. Swindon Zydis Limited have filed a patent application in 2019 (PCT/EP2019/082940—Oral dispersible vaccine comprising virosomes) with inventors D.W., C.S., M. Amacker, S.F., and T.S. that are authors of this manuscript. The remaining authors declare no competing interests.
Figures
Similar articles
-
A randomized placebo-controlled phase Ia malaria vaccine trial of two virosome-formulated synthetic peptides in healthy adult volunteers.PLoS One. 2007 Oct 10;2(10):e1018. doi: 10.1371/journal.pone.0001018. PLoS One. 2007. PMID: 17925866 Free PMC article. Clinical Trial.
-
The virosome concept for influenza vaccines.Vaccine. 2005 Jul 8;23 Suppl 1:S26-38. doi: 10.1016/j.vaccine.2005.04.026. Vaccine. 2005. PMID: 16026906 Review.
-
Evaluation of an intranasal virosomal vaccine against respiratory syncytial virus in mice: effect of TLR2 and NOD2 ligands on induction of systemic and mucosal immune responses.PLoS One. 2013 Apr 8;8(4):e61287. doi: 10.1371/journal.pone.0061287. Print 2013. PLoS One. 2013. PMID: 23593453 Free PMC article.
-
Influenza virosome/DNA vaccine complex as a new formulation to induce intra-subtypic protection against influenza virus challenge.Antiviral Res. 2012 Sep;95(3):229-36. doi: 10.1016/j.antiviral.2012.07.003. Epub 2012 Jul 15. Antiviral Res. 2012. PMID: 22809863
-
Influenza vaccines: the virosome concept.Immunol Lett. 2009 Feb 21;122(2):118-21. doi: 10.1016/j.imlet.2008.11.006. Epub 2008 Dec 25. Immunol Lett. 2009. PMID: 19100779 Review.
Cited by
-
Development of thermostable vaccine adjuvants.Expert Rev Vaccines. 2021 May;20(5):497-517. doi: 10.1080/14760584.2021.1902314. Epub 2021 Jun 26. Expert Rev Vaccines. 2021. PMID: 33724133 Free PMC article. Review.
-
Clinical and Preclinical Methods of Heat-Stabilization of Human Vaccines.Mol Pharm. 2024 Mar 4;21(3):1015-1026. doi: 10.1021/acs.molpharmaceut.3c00844. Epub 2024 Jan 30. Mol Pharm. 2024. PMID: 38288698 Free PMC article. Review.
-
Immunogenicity of Recombinant Lipid-Based Nanoparticle Vaccines: Danger Signal vs. Helping Hand.Pharmaceutics. 2023 Dec 23;16(1):24. doi: 10.3390/pharmaceutics16010024. Pharmaceutics. 2023. PMID: 38258035 Free PMC article. Review.
-
Recombinant protein vaccines, a proven approach against coronavirus pandemics.Adv Drug Deliv Rev. 2021 Mar;170:71-82. doi: 10.1016/j.addr.2021.01.001. Epub 2021 Jan 7. Adv Drug Deliv Rev. 2021. PMID: 33421475 Free PMC article. Review.
-
A spike virosome vaccine induces pan-sarbecovirus antibody responses in mice.iScience. 2024 Apr 10;27(5):109719. doi: 10.1016/j.isci.2024.109719. eCollection 2024 May 17. iScience. 2024. PMID: 38706848 Free PMC article.
References
-
- Chen D, Kristensen D. Opportunities and challenges of developing thermostable vaccines. Expert Rev. Vaccines. 2009;8:547–557. - PubMed
-
- de Jonge J, et al. Inulin sugar glasses preserve the structural integrity and biological activity of influenza virosomes during freeze-drying and storage. Eur. J. Pharm. Sci. 2007;32:33–44. - PubMed
-
- Karp CL, et al. Evaluating the value proposition for improving vaccine thermostability to increase vaccine impact in low and middle-income countries. Vaccine. 2015;33:3471–3479. - PubMed
-
- Kristensen D, Chen D, Cummings R. Vaccine stabilization: research, commercialization, and potential impact. Vaccine. 2011;29:7122–7124. - PubMed
-
- Lloyd J, Cheyne J. The origins of the vaccine cold chain and a glimpse of the future. Vaccine. 2017;35:2115–2120. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
