

A Circuit Encoding Absolute Cold Temperature in Drosophila
- PMID: 32442464
- PMCID: PMC7314653
- DOI: 10.1016/j.cub.2020.04.038
A Circuit Encoding Absolute Cold Temperature in Drosophila
Abstract
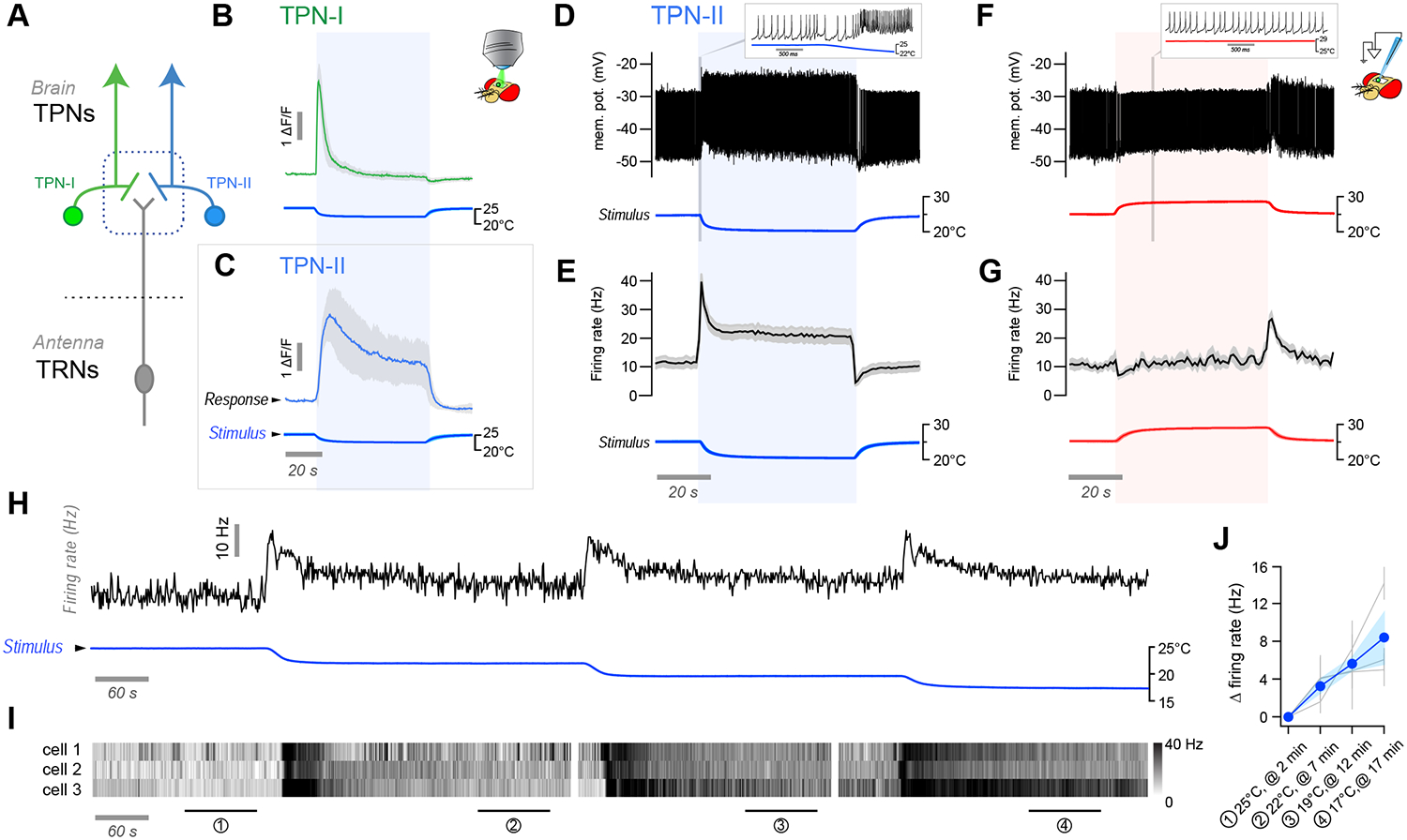
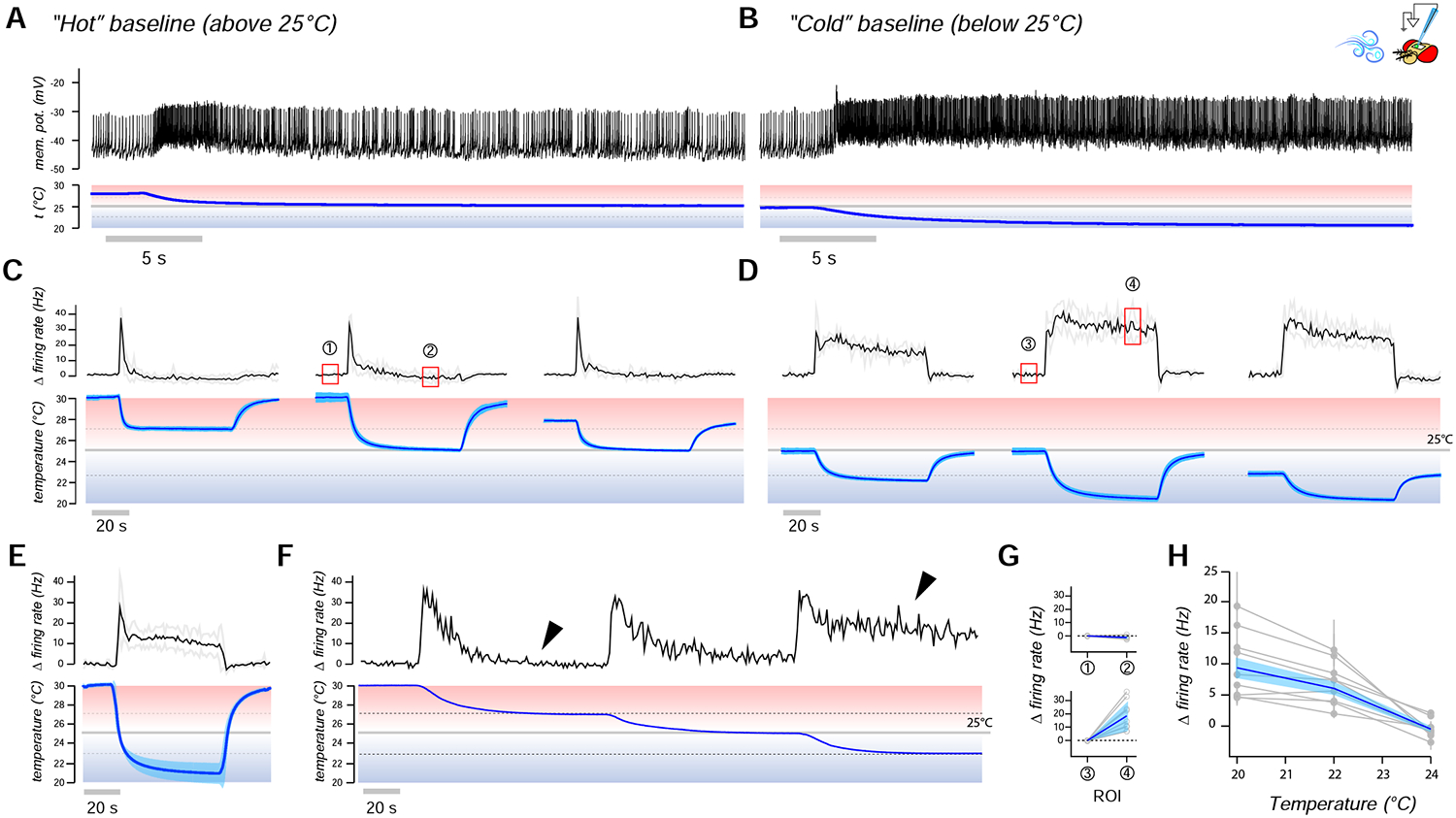
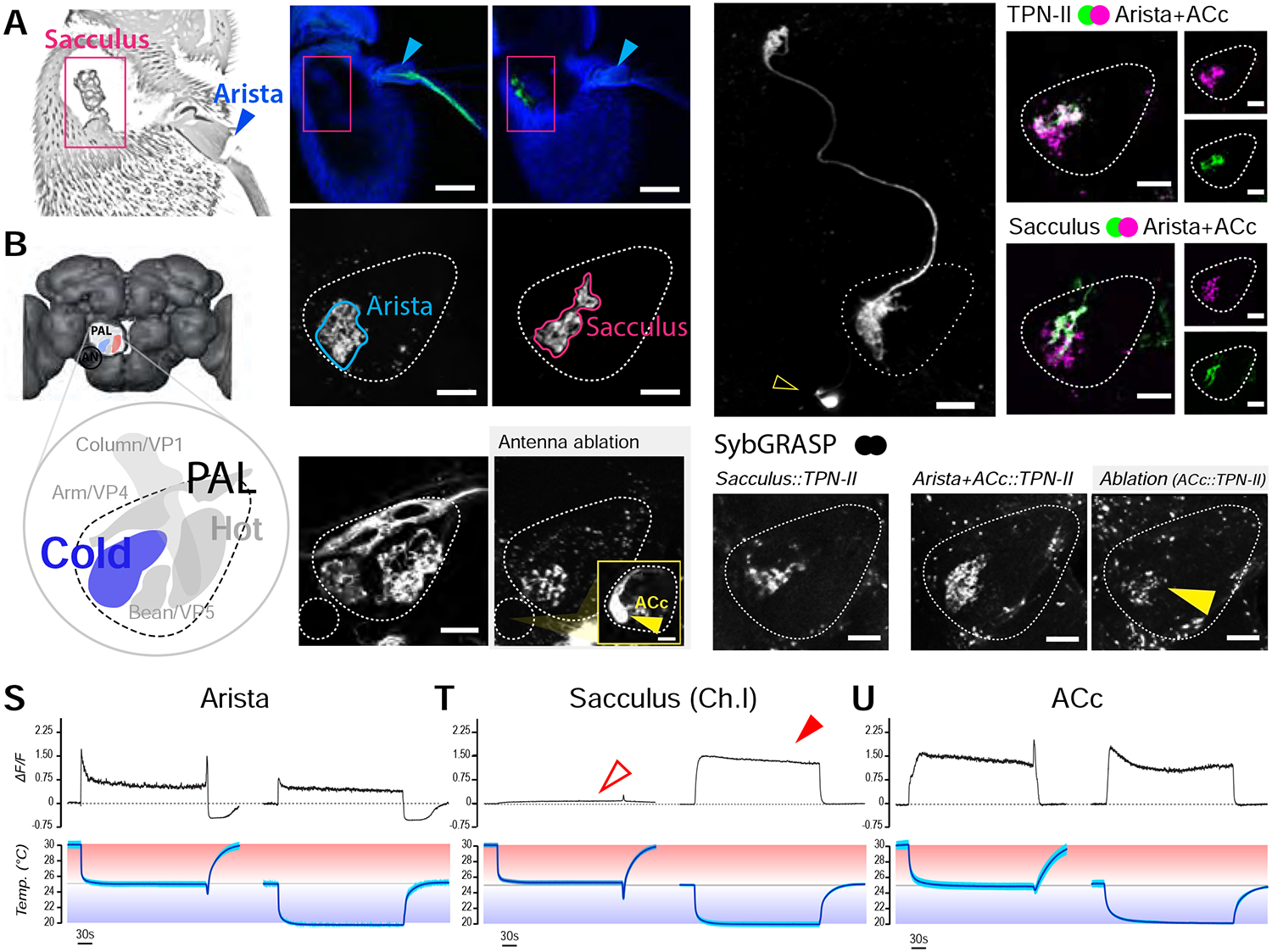
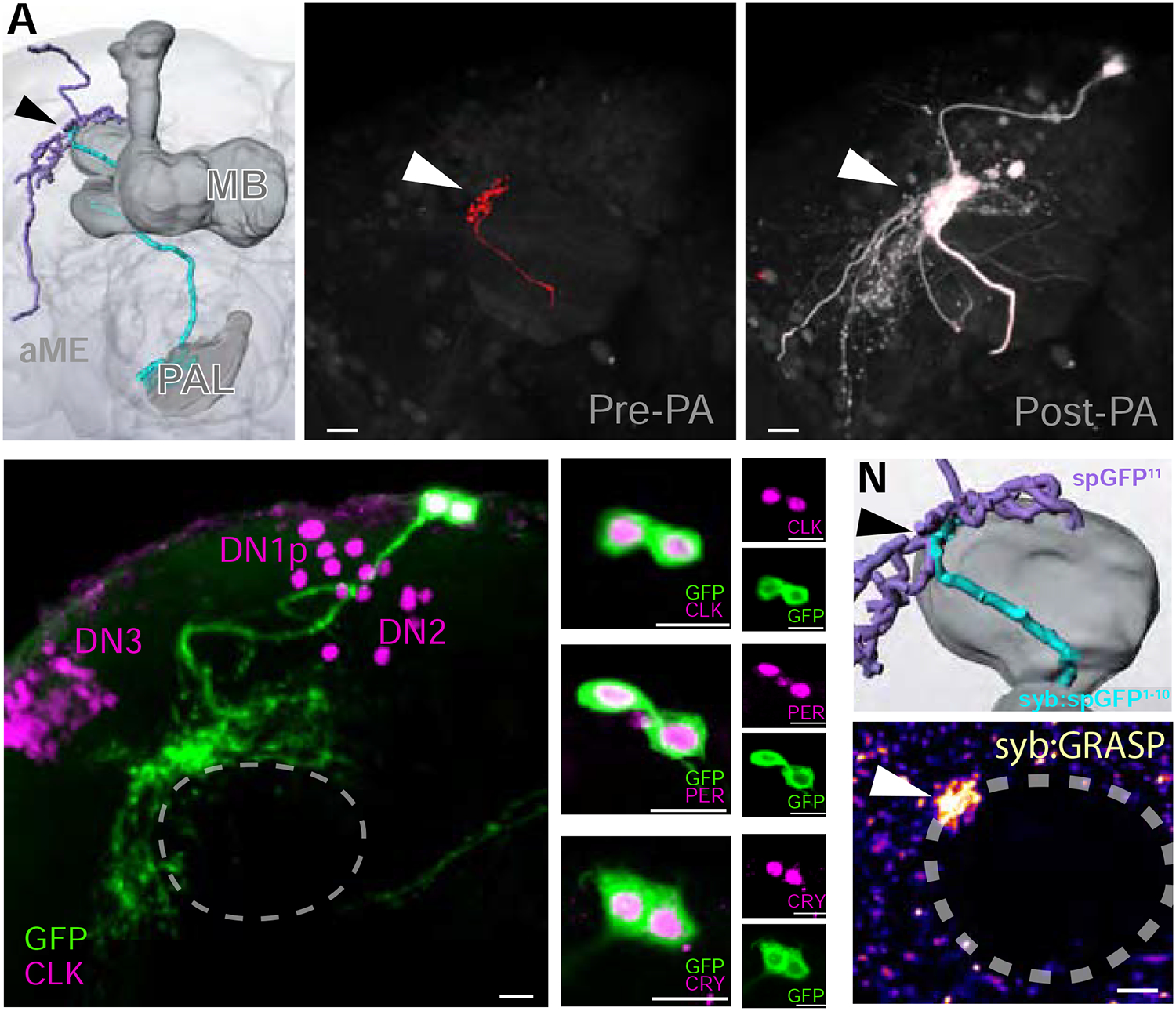
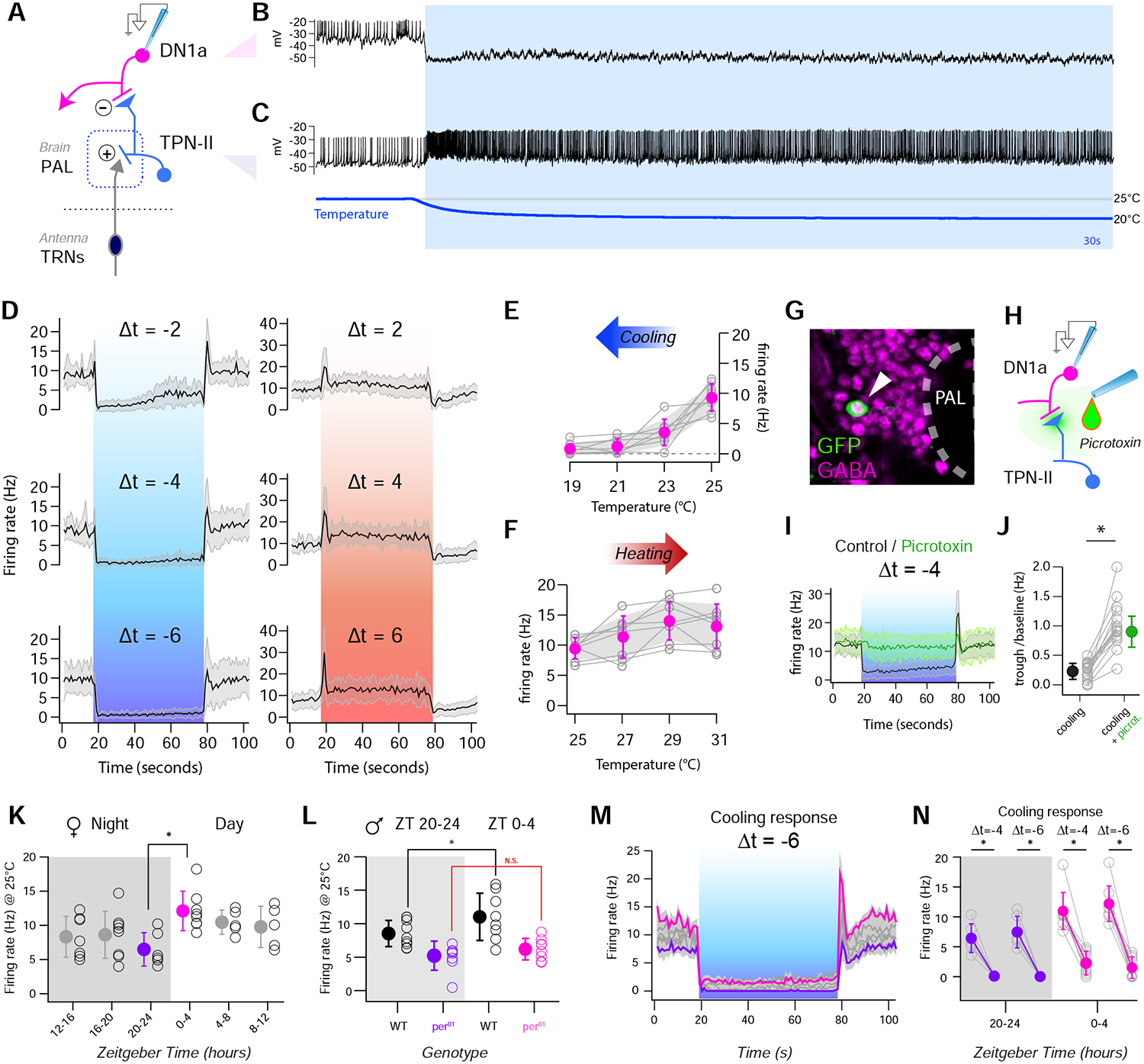
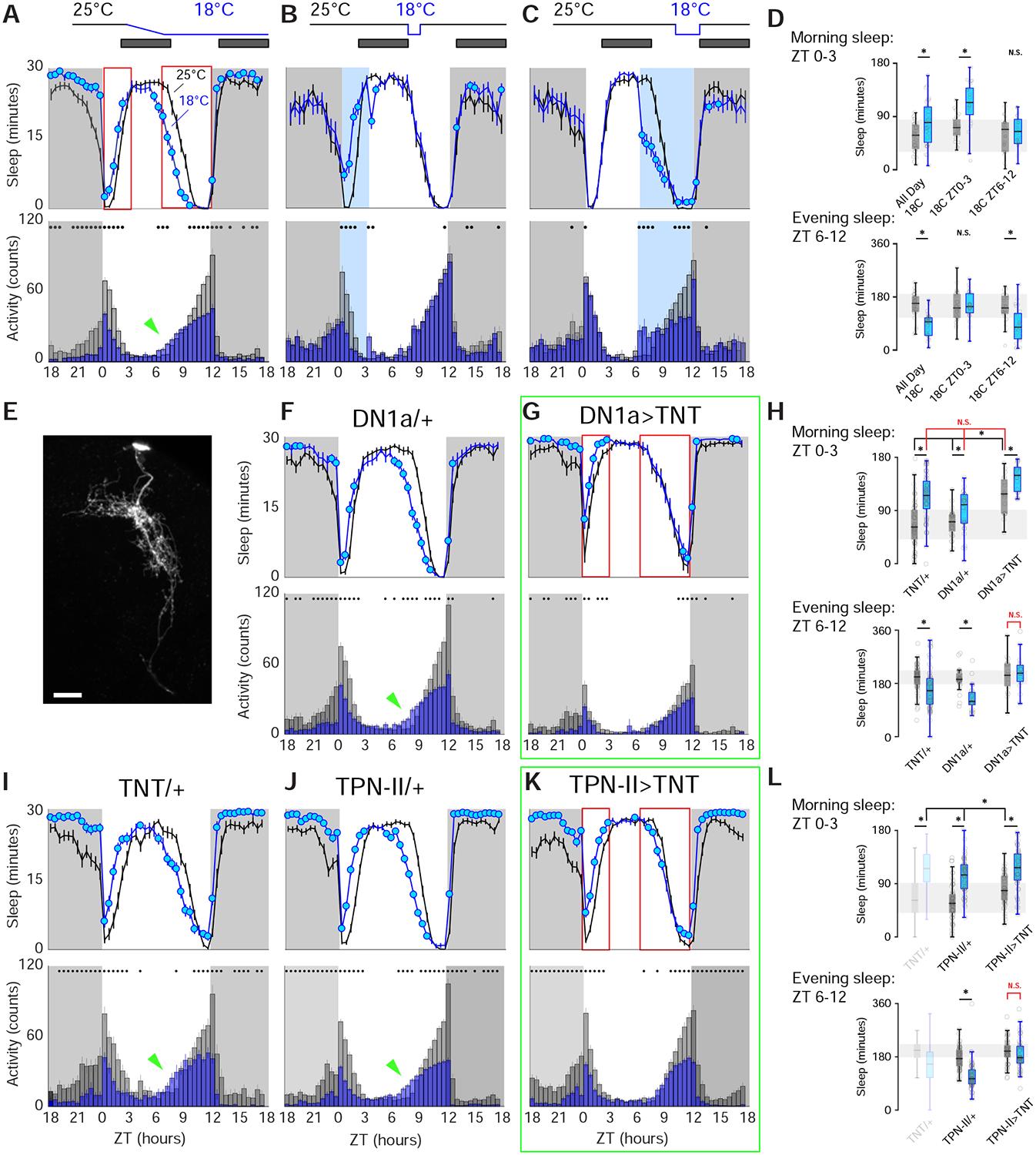
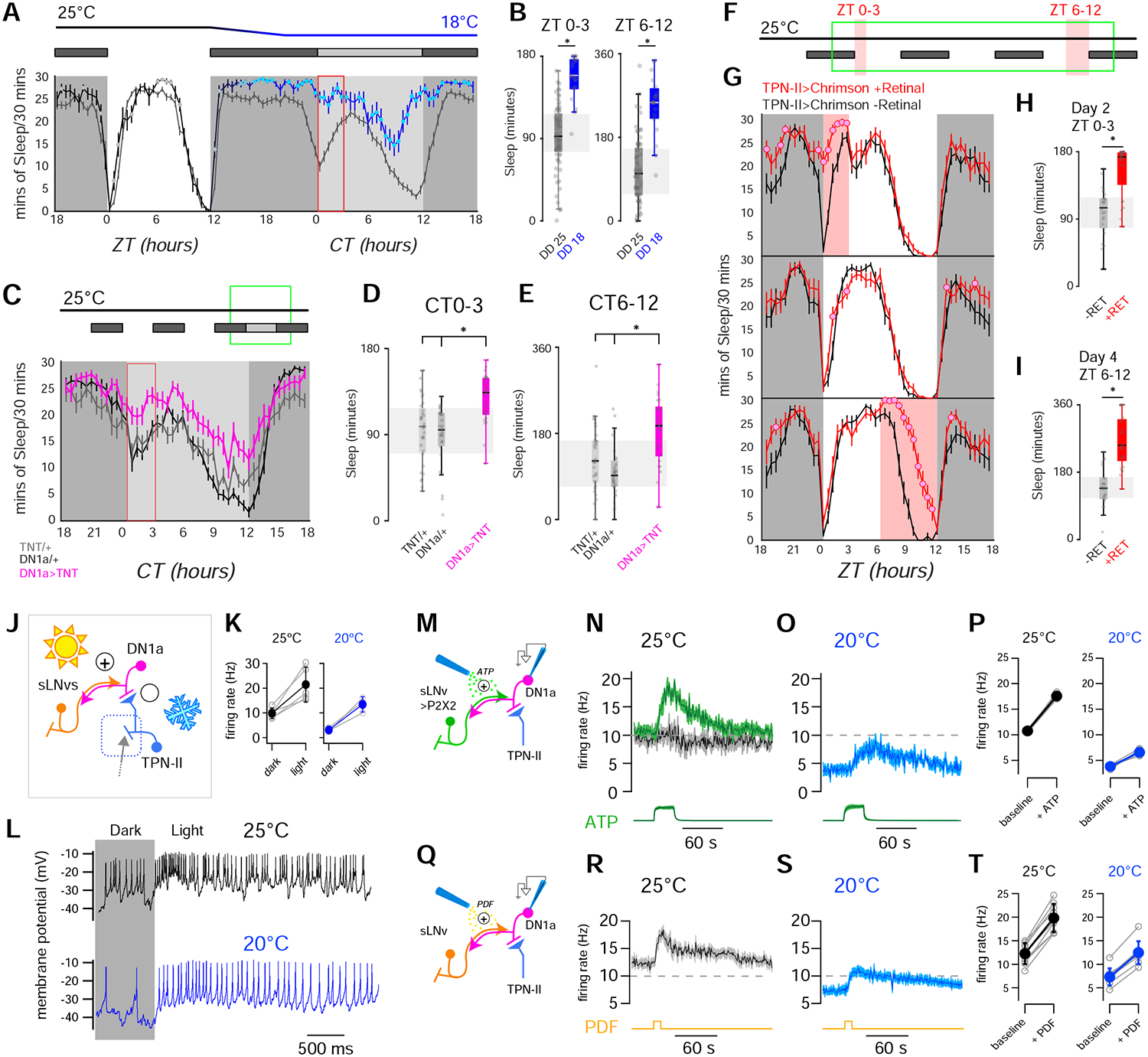
Animals react to environmental changes over timescales ranging from seconds to days and weeks. An important question is how sensory stimuli are parsed into neural signals operating over such diverse temporal scales. Here, we uncover a specialized circuit, from sensory neurons to higher brain centers, that processes information about long-lasting, absolute cold temperature in Drosophila. We identify second-order thermosensory projection neurons (TPN-IIs) exhibiting sustained firing that scales with absolute temperature. Strikingly, this activity only appears below the species-specific, preferred temperature for D. melanogaster (∼25°C). We trace the inputs and outputs of TPN-IIs and find that they are embedded in a cold "thermometer" circuit that provides powerful and persistent inhibition to brain centers involved in regulating sleep and activity. Our results demonstrate that the fly nervous system selectively encodes and relays absolute temperature information and illustrate a sensory mechanism that allows animals to adapt behavior specifically to cold conditions on the timescale of hours to days.
Keywords: Drosophila; antenna; brain; circuit; cold; seasonal; sleep; temperature; thermosensory.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
Comment in
-
Neuroscience: Sensing Absolute Cold.Curr Biol. 2020 Jul 20;30(14):R809-R811. doi: 10.1016/j.cub.2020.05.026. Curr Biol. 2020. PMID: 32693074 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
