

Posttranslational Regulation of the Exon Skipping Machinery Controls Aberrant Splicing in Leukemia
- PMID: 32444465
- PMCID: PMC7483384
- DOI: 10.1158/2159-8290.CD-19-1436
Posttranslational Regulation of the Exon Skipping Machinery Controls Aberrant Splicing in Leukemia
Abstract
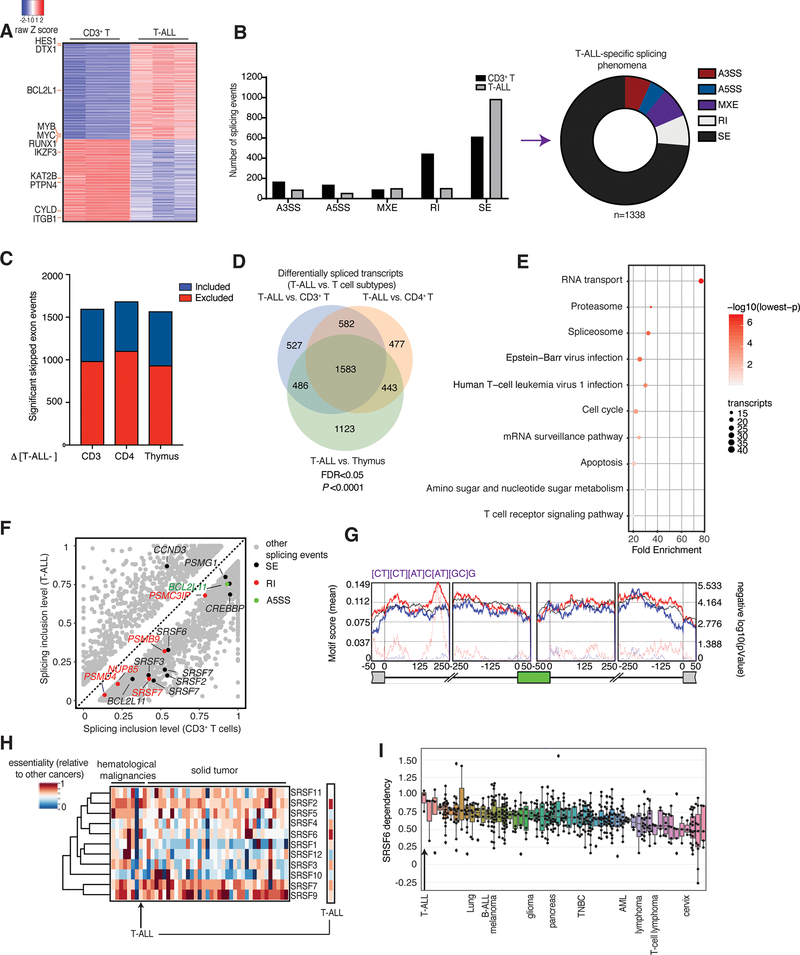
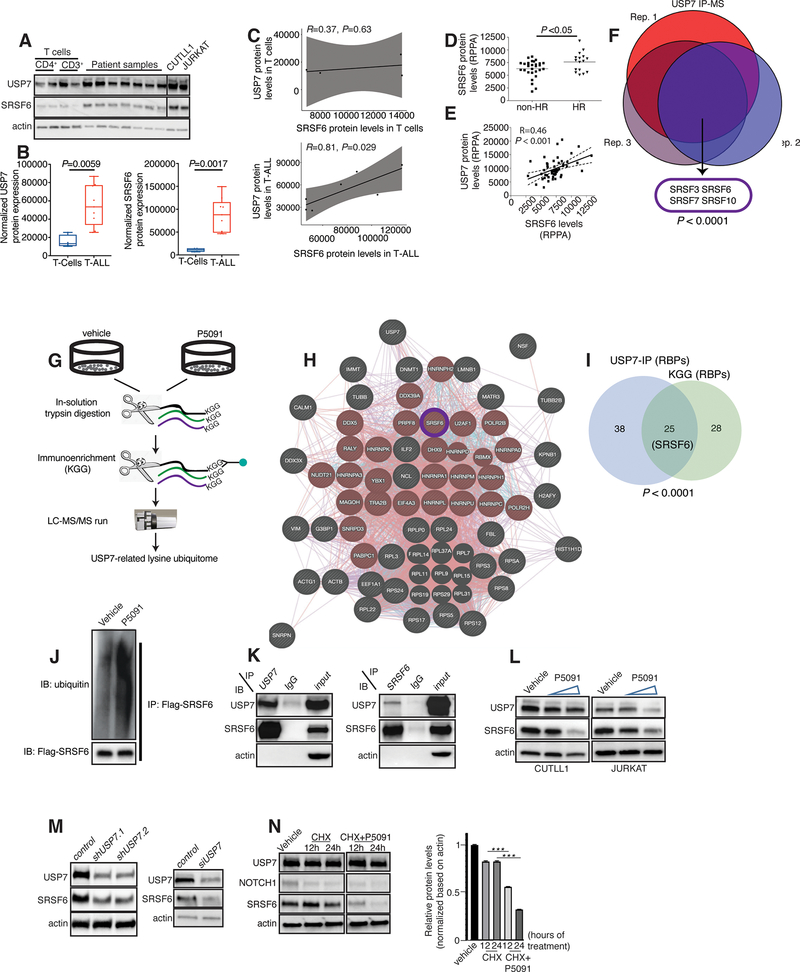
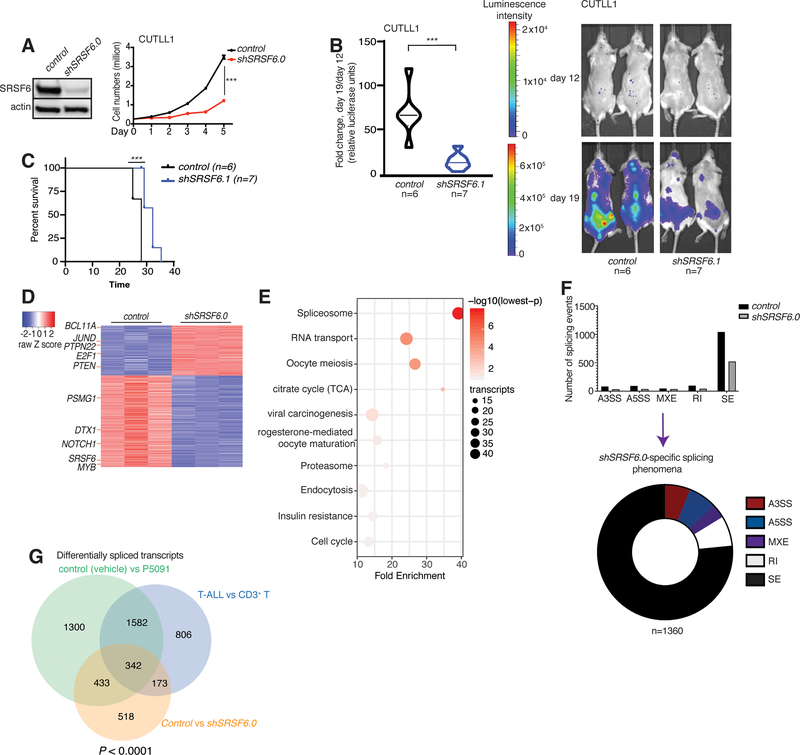
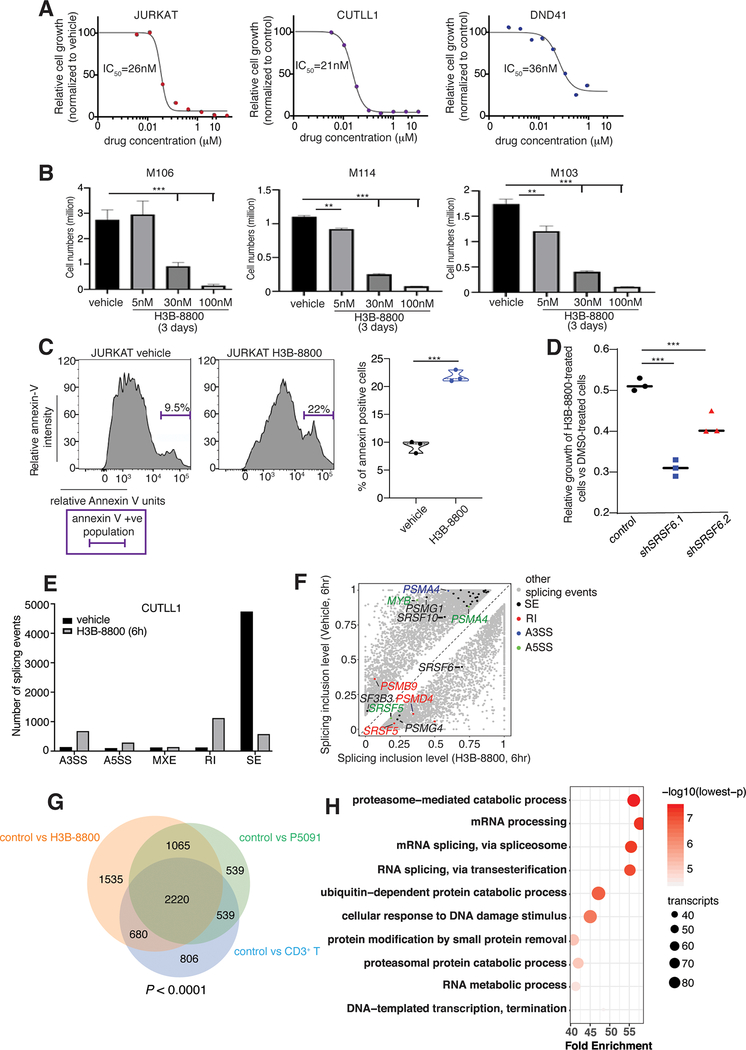
Splicing alterations are common in diseases such as cancer, where mutations in splicing factor genes are frequently responsible for aberrant splicing. Here we present an alternative mechanism for splicing regulation in T-cell acute lymphoblastic leukemia (T-ALL) that involves posttranslational stabilization of the splicing machinery via deubiquitination. We demonstrate there are extensive exon skipping changes in disease, affecting proteasomal subunits, cell-cycle regulators, and the RNA machinery. We present that the serine/arginine-rich splicing factors (SRSF), controlling exon skipping, are critical for leukemia cell survival. The ubiquitin-specific peptidase 7 (USP7) regulates SRSF6 protein levels via active deubiquitination, and USP7 inhibition alters the exon skipping pattern and blocks T-ALL growth. The splicing inhibitor H3B-8800 affects splicing of proteasomal transcripts and proteasome activity and acts synergistically with proteasome inhibitors in inhibiting T-ALL growth. Our study provides the proof-of-principle for regulation of splicing factors via deubiquitination and suggests new therapeutic modalities in T-ALL. SIGNIFICANCE: Our study provides a new proof-of-principle for posttranslational regulation of splicing factors independently of mutations in aggressive T-cell leukemia. It further suggests a new drug combination of splicing and proteasomal inhibitors, a concept that might apply to other diseases with or without mutations affecting the splicing machinery.This article is highlighted in the In This Issue feature, p. 1241.
©2020 American Association for Cancer Research.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- P30 CA016087/CA/NCI NIH HHS/United States
- T32 CA009560/CA/NCI NIH HHS/United States
- R01 CA228135/CA/NCI NIH HHS/United States
- P41 GM108569/GM/NIGMS NIH HHS/United States
- P30 CA060553/CA/NCI NIH HHS/United States
- R01 CA216421/CA/NCI NIH HHS/United States
- S10 OD023423/OD/NIH HHS/United States
- R00 CA175293/CA/NCI NIH HHS/United States
- T32 GM144295/GM/NIGMS NIH HHS/United States
- R01 CA248770/CA/NCI NIH HHS/United States
- P01 CA229086/CA/NCI NIH HHS/United States
- T32 GM008152/GM/NIGMS NIH HHS/United States
- S10 OD025194/OD/NIH HHS/United States
- R00 CA188293/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
