

Truncation of mutant huntingtin in knock-in mice demonstrates exon1 huntingtin is a key pathogenic form
- PMID: 32444599
- PMCID: PMC7244548
- DOI: 10.1038/s41467-020-16318-1
Truncation of mutant huntingtin in knock-in mice demonstrates exon1 huntingtin is a key pathogenic form
Erratum in
-
Author Correction: Truncation of mutant huntingtin in knock-in mice demonstrates exon1 huntingtin is a key pathogenic form.Nat Commun. 2020 Nov 19;11(1):5989. doi: 10.1038/s41467-020-19873-9. Nat Commun. 2020. PMID: 33214567 Free PMC article.
Abstract
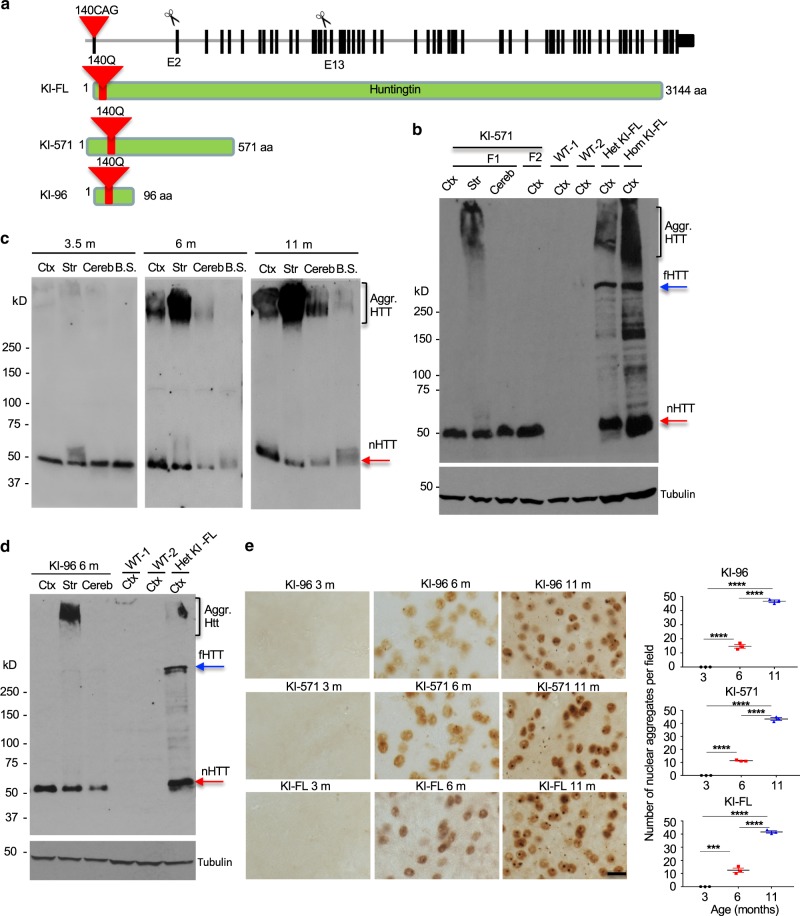
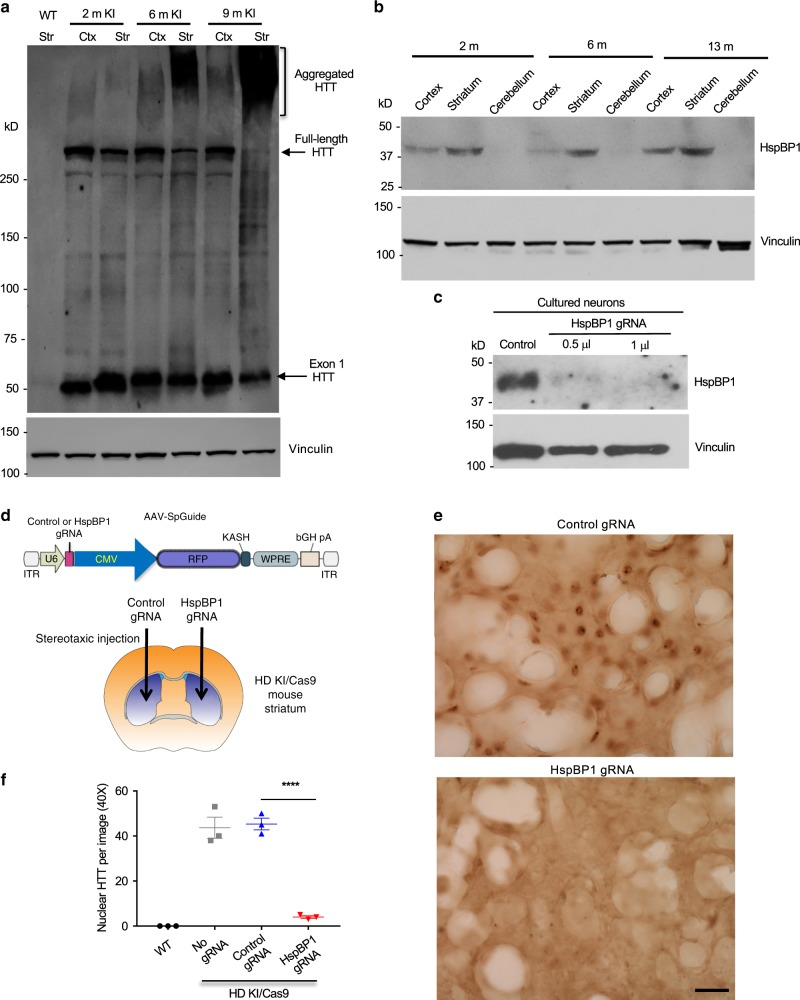
Polyglutamine expansion in proteins can cause selective neurodegeneration, although the mechanisms are not fully understood. In Huntington's disease (HD), proteolytic processing generates toxic N-terminal huntingtin (HTT) fragments that preferentially kill striatal neurons. Here, using CRISPR/Cas9 to truncate full-length mutant HTT in HD140Q knock-in (KI) mice, we show that exon 1 HTT is stably present in the brain, regardless of truncation sites in full-length HTT. This N-terminal HTT leads to similar HD-like phenotypes and age-dependent HTT accumulation in the striatum in different KI mice. We find that exon 1 HTT is constantly generated but its selective accumulation in the striatum is associated with the age-dependent expression of striatum-enriched HspBP1, a chaperone inhibitory protein. Our findings suggest that tissue-specific chaperone function contributes to the selective neuropathology in HD, and highlight the therapeutic potential in blocking generation of exon 1 HTT.
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Orr HT, Zoghbi HY. Trinucleotide repeat disorders. Annu Rev. Neurosci. 2007;30:575–621. - PubMed
-
- Bates GP, et al. Huntington disease. Nat. Rev. Dis. Prim. 2015;1:15005. - PubMed
-
- Ross CA, Tabrizi SJ. Huntington’s disease: from molecular pathogenesis to clinical treatment. Lancet Neurol. 2011;10:83–98. - PubMed
-
- Vonsattel JP, et al. Neuropathological classification of Huntington’s disease. J. Neuropathol. Exp. Neurol. 1985;44:559–577. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
