

An integrated multi-omics approach identifies the landscape of interferon-α-mediated responses of human pancreatic beta cells
- PMID: 32444635
- PMCID: PMC7244579
- DOI: 10.1038/s41467-020-16327-0
An integrated multi-omics approach identifies the landscape of interferon-α-mediated responses of human pancreatic beta cells
Abstract
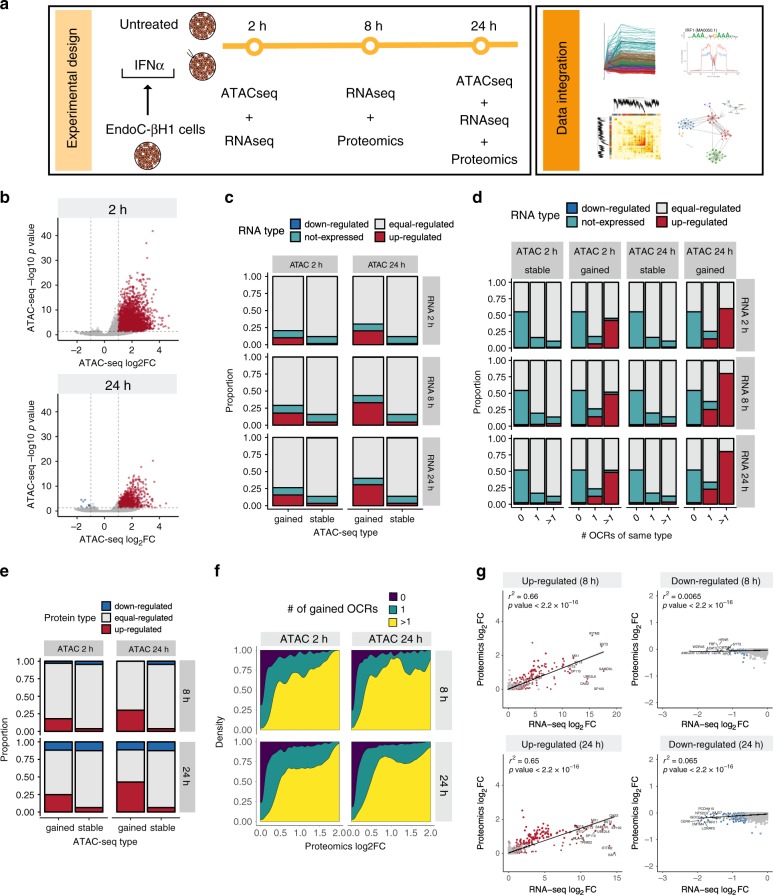
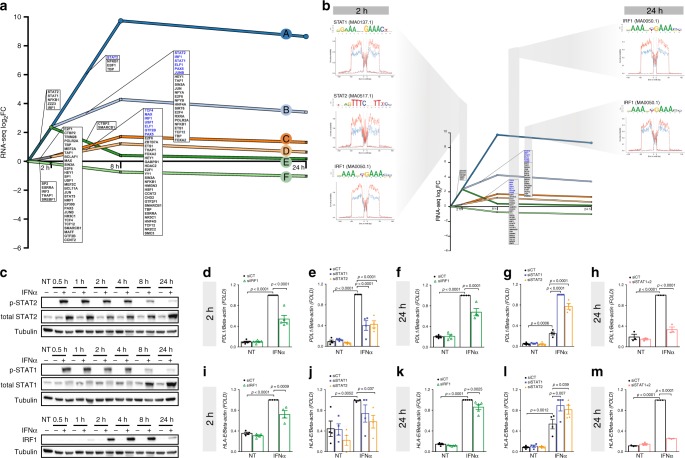
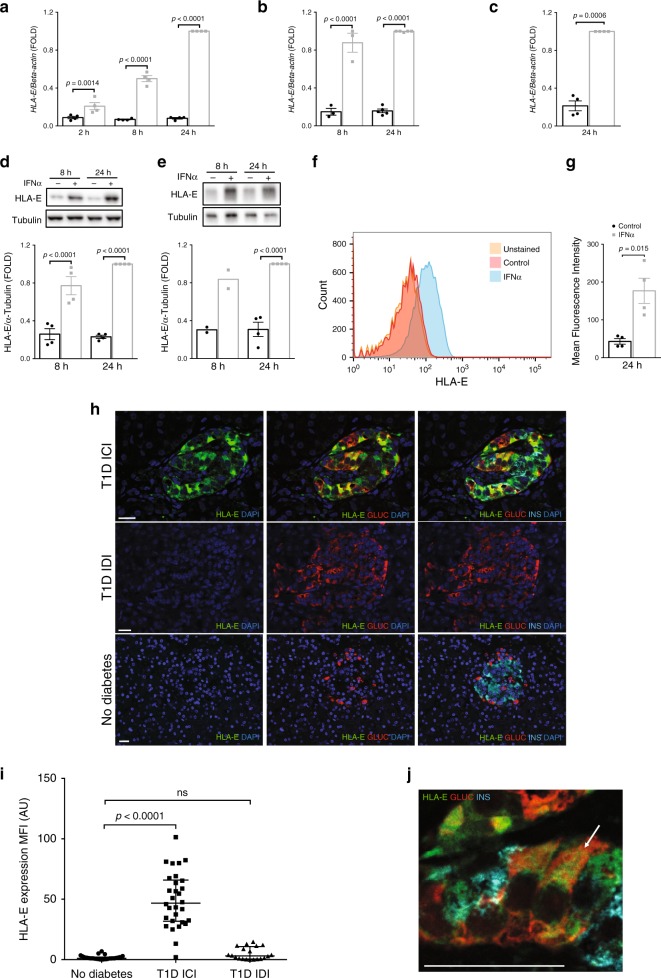
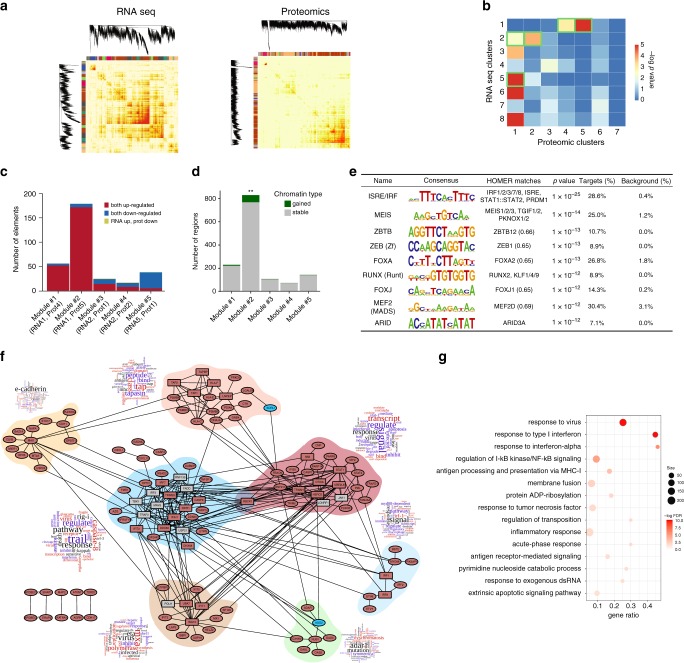
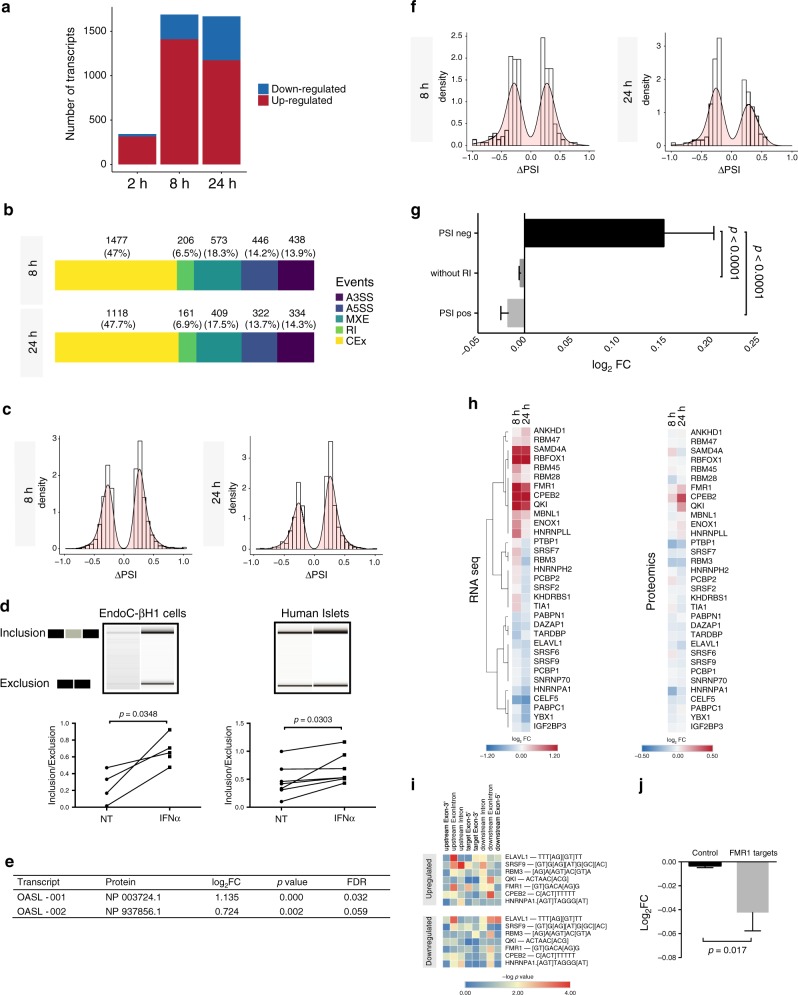
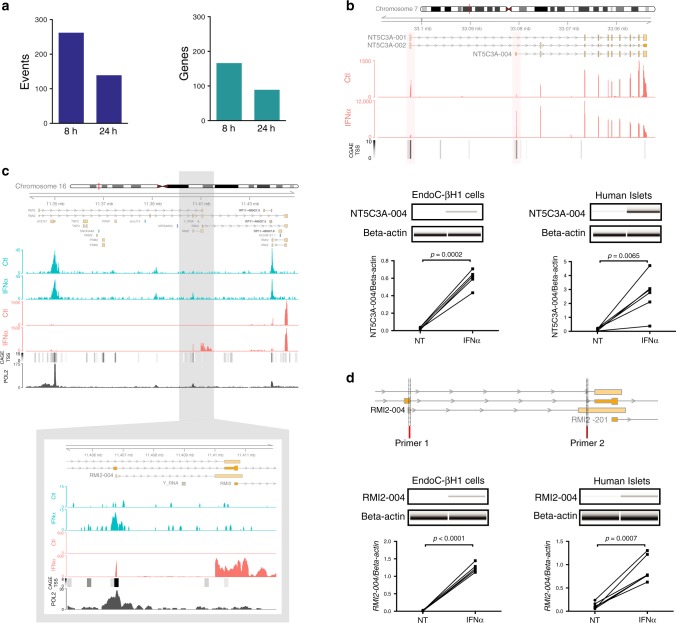
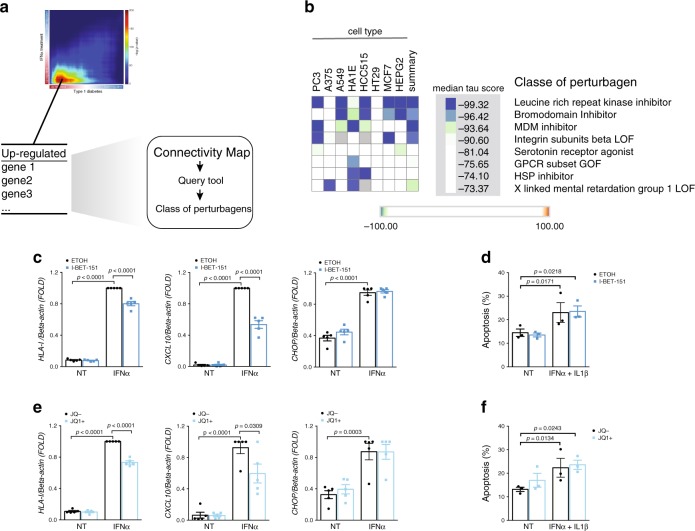
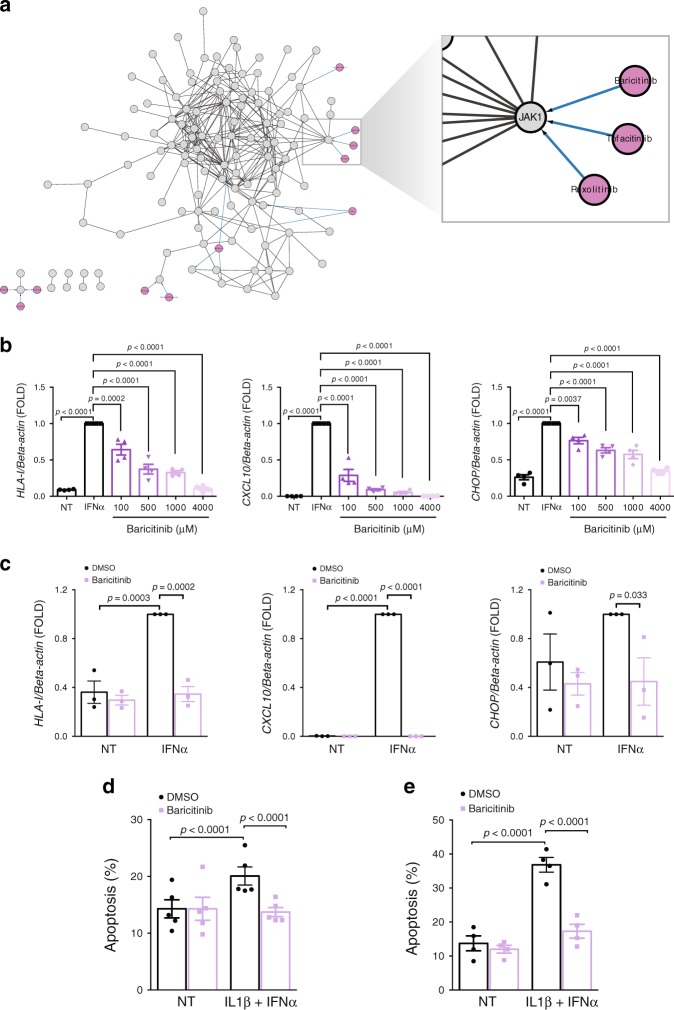
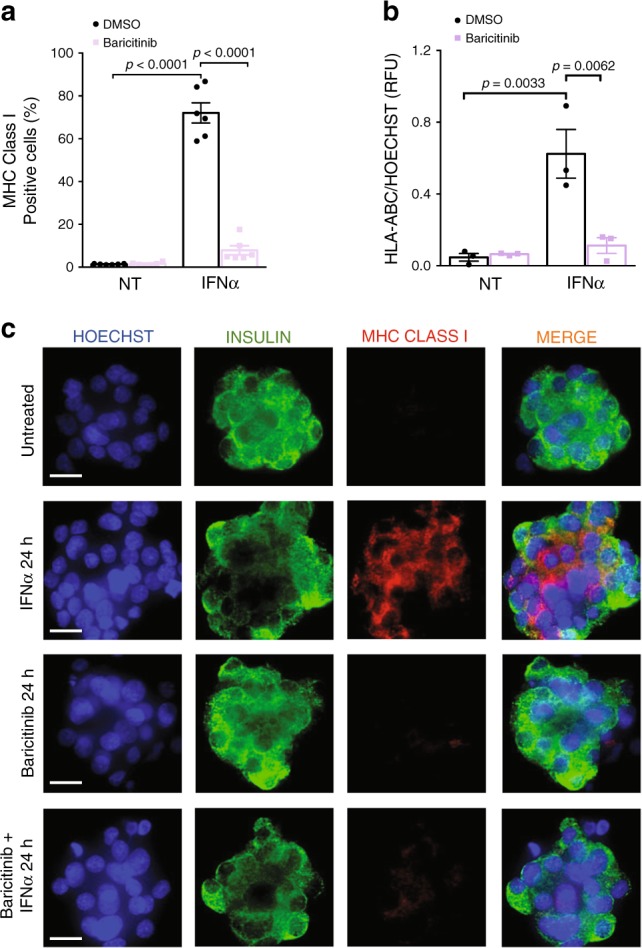
Interferon-α (IFNα), a type I interferon, is expressed in the islets of type 1 diabetic individuals, and its expression and signaling are regulated by T1D genetic risk variants and viral infections associated with T1D. We presently characterize human beta cell responses to IFNα by combining ATAC-seq, RNA-seq and proteomics assays. The initial response to IFNα is characterized by chromatin remodeling, followed by changes in transcriptional and translational regulation. IFNα induces changes in alternative splicing (AS) and first exon usage, increasing the diversity of transcripts expressed by the beta cells. This, combined with changes observed on protein modification/degradation, ER stress and MHC class I, may expand antigens presented by beta cells to the immune system. Beta cells also up-regulate the checkpoint proteins PDL1 and HLA-E that may exert a protective role against the autoimmune assault. Data mining of the present multi-omics analysis identifies two compound classes that antagonize IFNα effects on human beta cells.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Eizirik DL, Colli ML, Ortis F. The role of inflammation in insulitis and beta-cell loss in type 1 diabetes. Nat. Rev. Endocrinol. 2009;5:219–226. - PubMed
-
- Crow MK, Olferiev M, Kirou KA. Type I interferons in autoimmune disease. Annu Rev. Pathol. 2019;14:369–393. - PubMed
-
- Eizirik DL, Op de Beeck A. Coxsackievirus and type 1 diabetes mellitus: the Wolf’s footprints. Trends Endocrinol. Metab. 2018;29:137–139. - PubMed
-
- Foulis AK, Farquharson MA, Meager A. Immunoreactive alpha-interferon in insulin-secreting beta cells in type 1 diabetes mellitus. Lancet. 1987;2:1423–1427. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
