

Phosphoproteomics identifies dual-site phosphorylation in an extended basophilic motif regulating FILIP1-mediated degradation of filamin-C
- PMID: 32444788
- PMCID: PMC7244511
- DOI: 10.1038/s42003-020-0982-5
Phosphoproteomics identifies dual-site phosphorylation in an extended basophilic motif regulating FILIP1-mediated degradation of filamin-C
Abstract
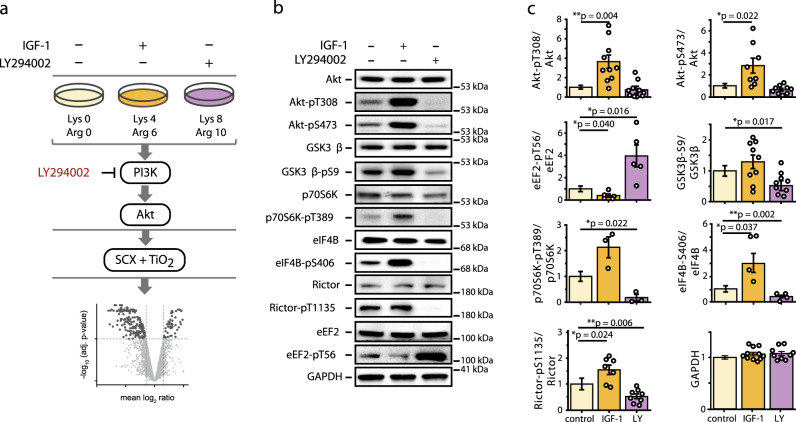
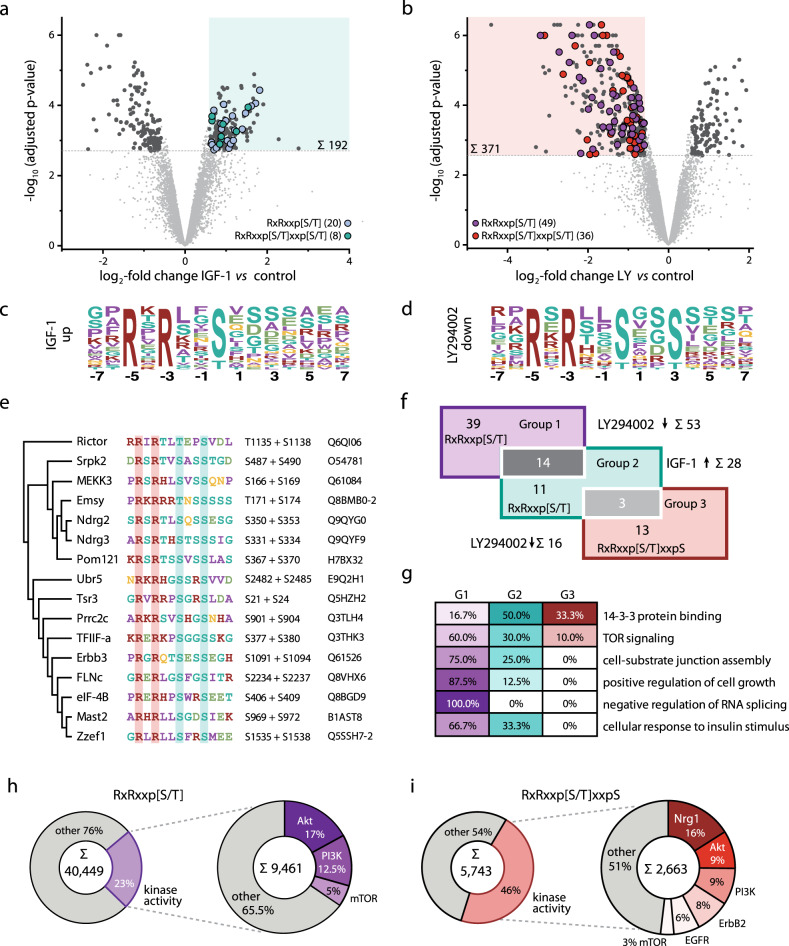
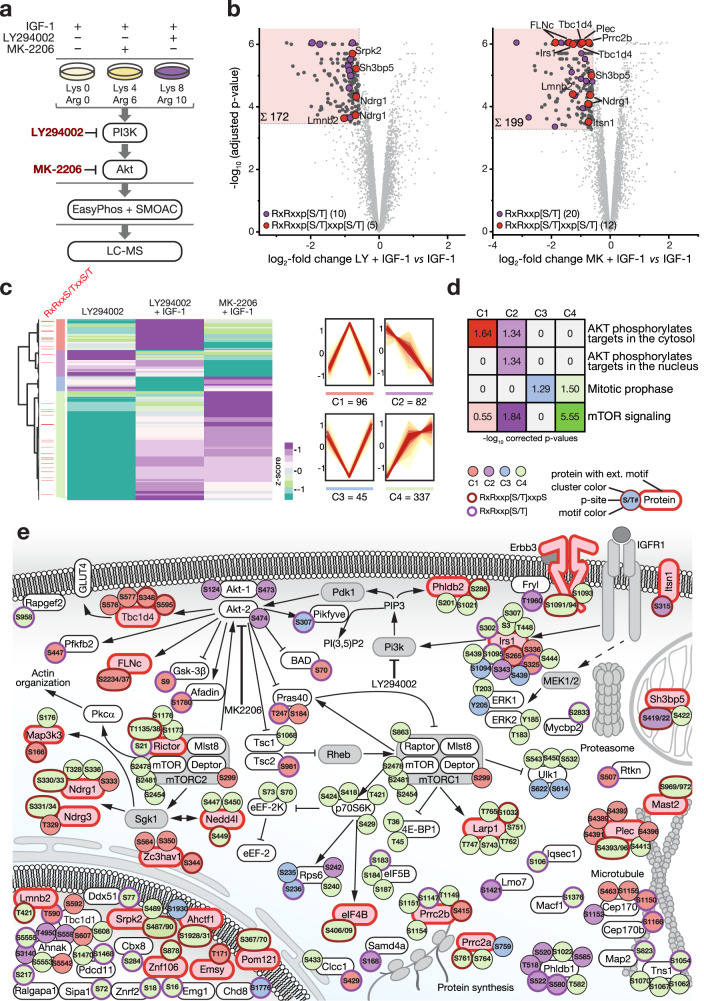
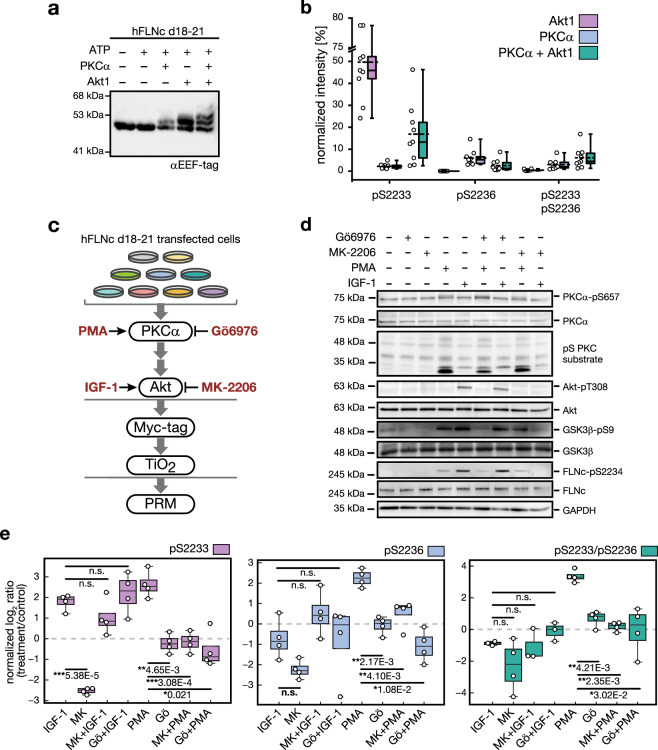
The PI3K/Akt pathway promotes skeletal muscle growth and myogenic differentiation. Although its importance in skeletal muscle biology is well documented, many of its substrates remain to be identified. We here studied PI3K/Akt signaling in contracting skeletal muscle cells by quantitative phosphoproteomics. We identified the extended basophilic phosphosite motif RxRxxp[S/T]xxp[S/T] in various proteins including filamin-C (FLNc). Importantly, this extended motif, located in a unique insert in Ig-like domain 20 of FLNc, is doubly phosphorylated. The protein kinases responsible for this dual-site phosphorylation are Akt and PKCα. Proximity proteomics and interaction analysis identified filamin A-interacting protein 1 (FILIP1) as direct FLNc binding partner. FILIP1 binding induces filamin degradation, thereby negatively regulating its function. Here, dual-site phosphorylation of FLNc not only reduces FILIP1 binding, providing a mechanism to shield FLNc from FILIP1-mediated degradation, but also enables fast dynamics of FLNc necessary for its function as signaling adaptor in cross-striated muscle cells.
Conflict of interest statement
The authors declare no competing interests.
Figures
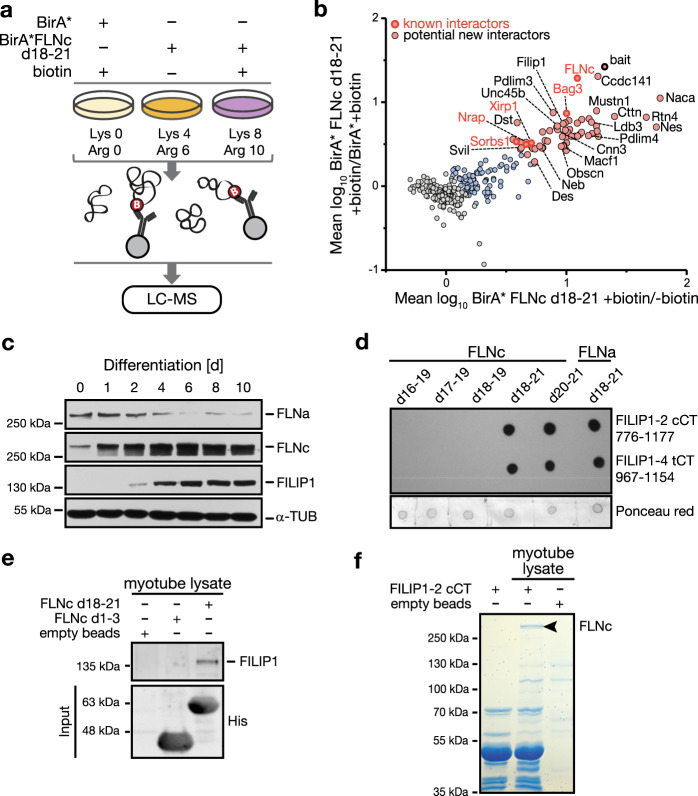
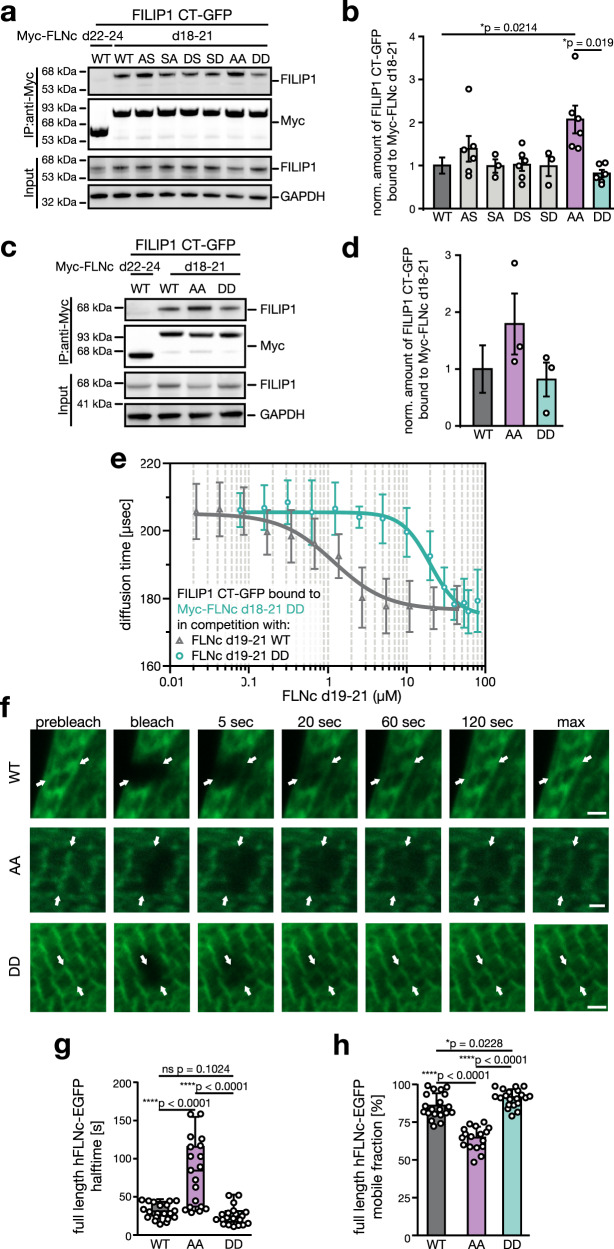
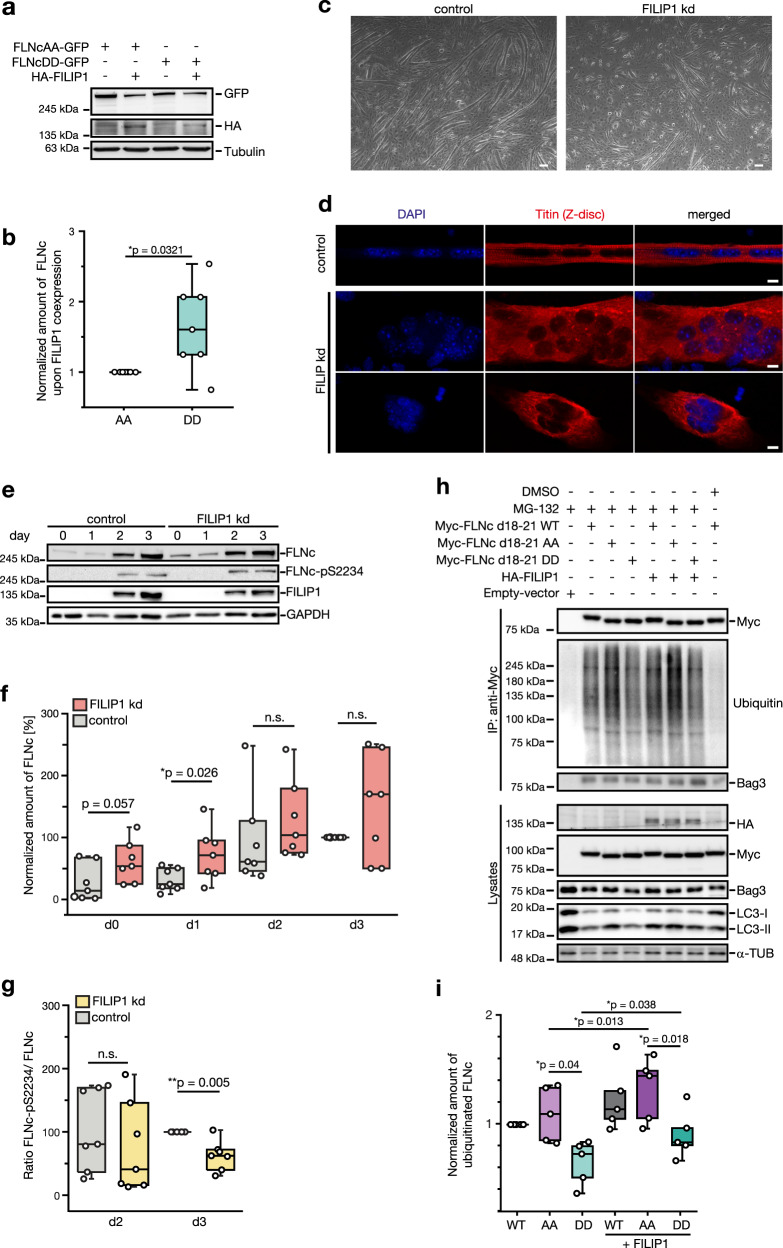
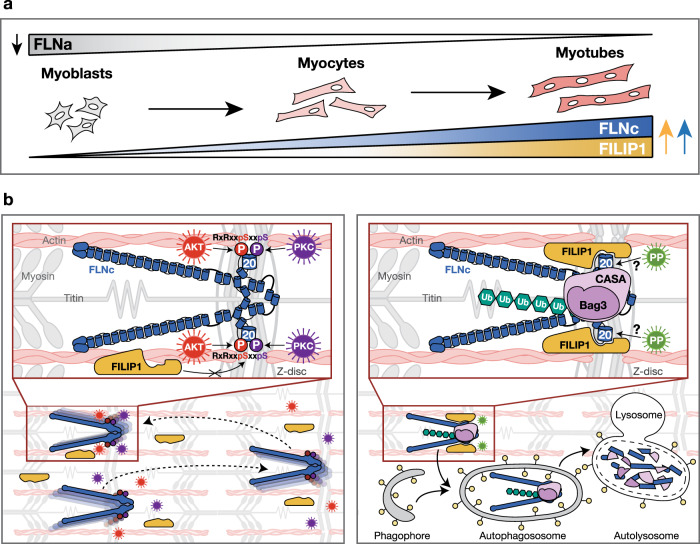
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
