

Metabolic Regulation of the Epigenome Drives Lethal Infantile Ependymoma
- PMID: 32445698
- PMCID: PMC10782558
- DOI: 10.1016/j.cell.2020.04.047
Metabolic Regulation of the Epigenome Drives Lethal Infantile Ependymoma
Abstract
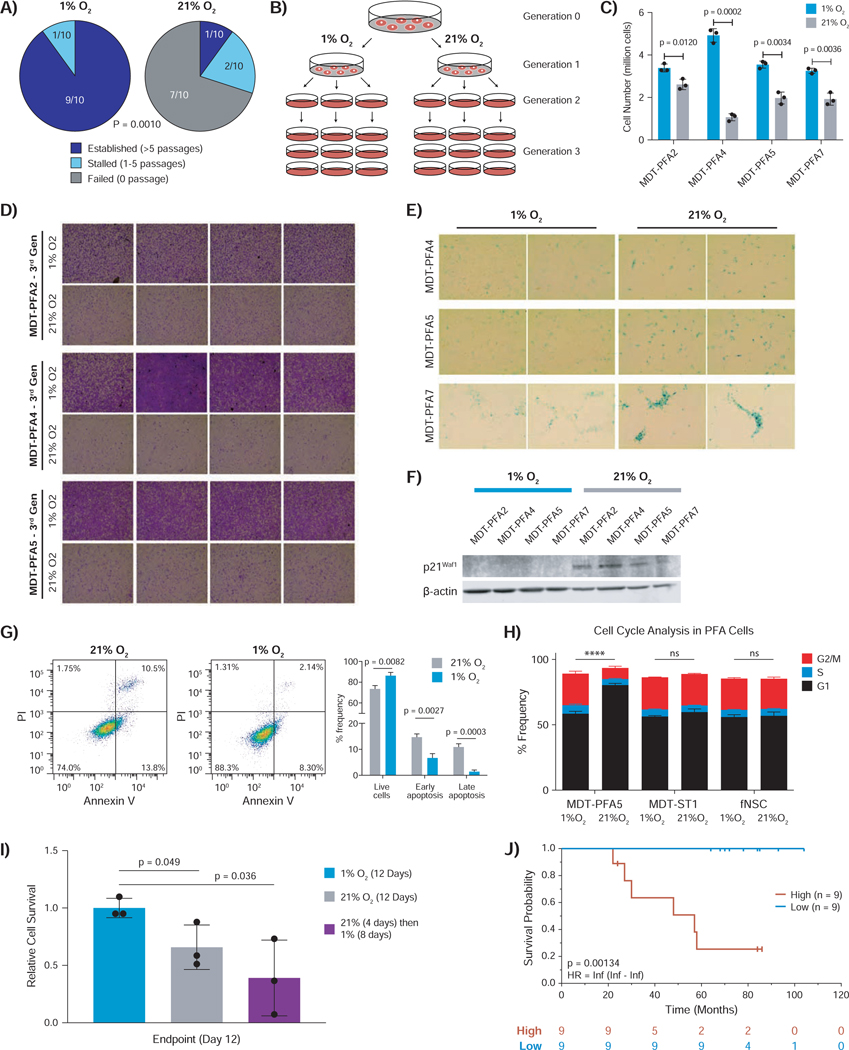
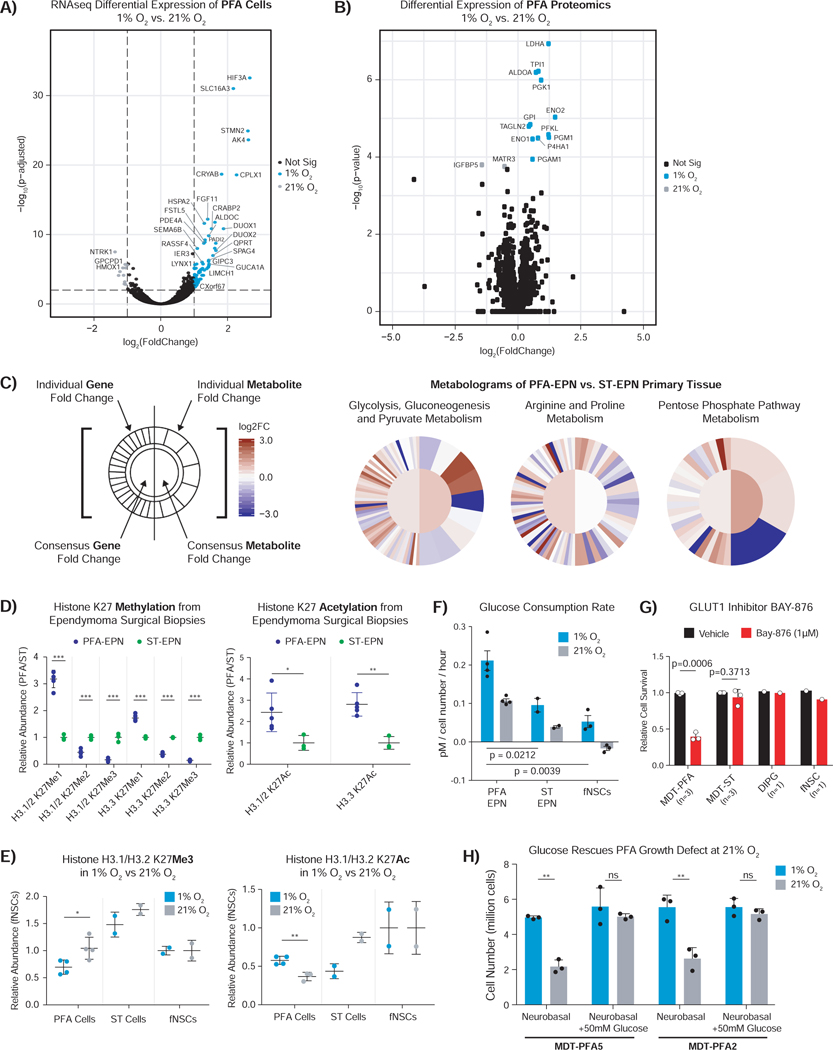
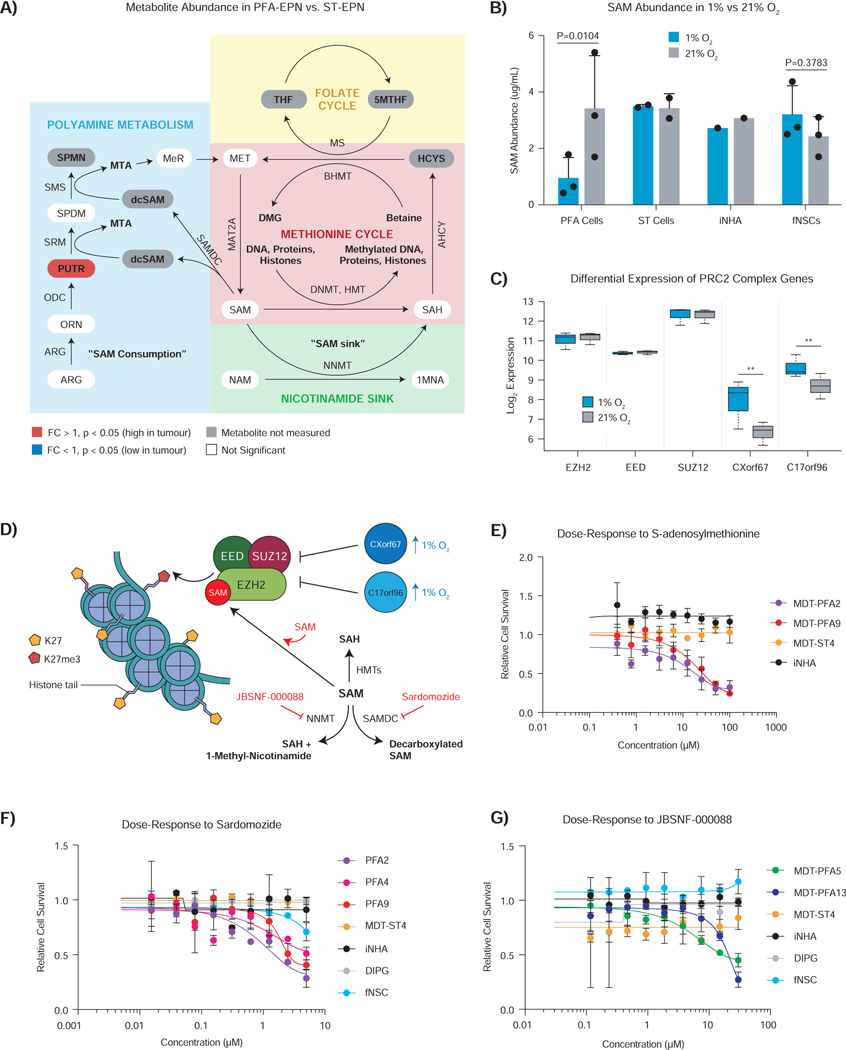
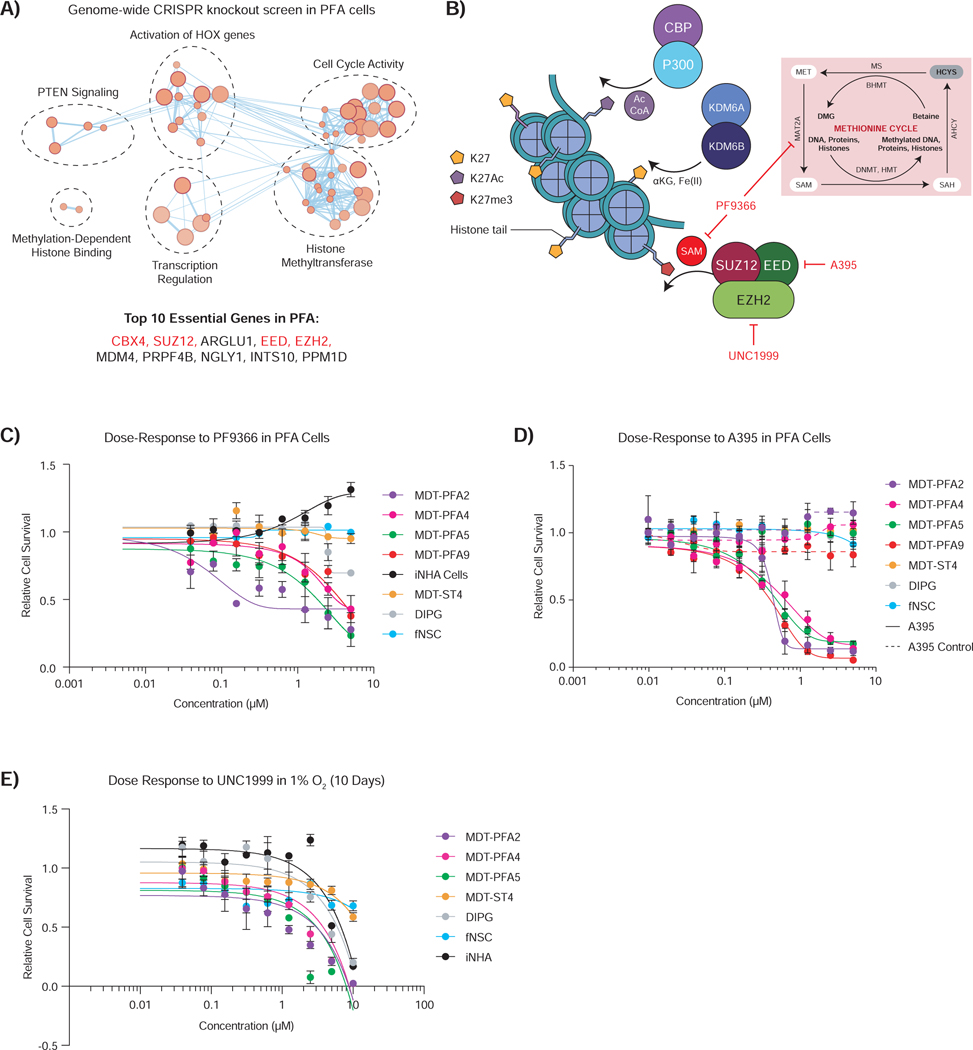
Posterior fossa A (PFA) ependymomas are lethal malignancies of the hindbrain in infants and toddlers. Lacking highly recurrent somatic mutations, PFA ependymomas are proposed to be epigenetically driven tumors for which model systems are lacking. Here we demonstrate that PFA ependymomas are maintained under hypoxia, associated with restricted availability of specific metabolites to diminish histone methylation, and increase histone demethylation and acetylation at histone 3 lysine 27 (H3K27). PFA ependymomas initiate from a cell lineage in the first trimester of human development that resides in restricted oxygen. Unlike other ependymomas, transient exposure of PFA cells to ambient oxygen induces irreversible cellular toxicity. PFA tumors exhibit a low basal level of H3K27me3, and, paradoxically, inhibition of H3K27 methylation specifically disrupts PFA tumor growth. Targeting metabolism and/or the epigenome presents a unique opportunity for rational therapy for infants with PFA ependymoma.
Keywords: cancer metabolism; ependymoma; epigenetics; hindbrain development; microenvironment; paediatric cancer.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures



Comment in
-
Understanding the Deadly Silence of Posterior Fossa A Ependymoma.Mol Cell. 2020 Jun 18;78(6):999-1001. doi: 10.1016/j.molcel.2020.05.020. Mol Cell. 2020. PMID: 32559429
-
Into Thin Air: Hypoxia Drives Metabolic and Epigenomic Deregulation of Lethal Pediatric Ependymoma.Dev Cell. 2020 Jul 20;54(2):134-136. doi: 10.1016/j.devcel.2020.06.020. Dev Cell. 2020. PMID: 32693052
References
-
- Andrews S. (2010). FastQC: a quality control tool for high throughput sequence data.
-
- Beringer M, Pisano P, Di Carlo V, Blanco E, Chammas P, Vizán P, Gutiérrez A, Aranda S, Payer B, Wierer M, et al. (2016). EPOP Functionally Links Elongin and Polycomb in Pluripotent Stem Cells. Mol. Cell 64, 645–658. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
