

Effect of extracellular vesicles from S. aureus-challenged human neutrophils on macrophages
- PMID: 32450612
- PMCID: PMC7686018
- DOI: 10.1002/JLB.3AB0320-156R
Effect of extracellular vesicles from S. aureus-challenged human neutrophils on macrophages
Abstract
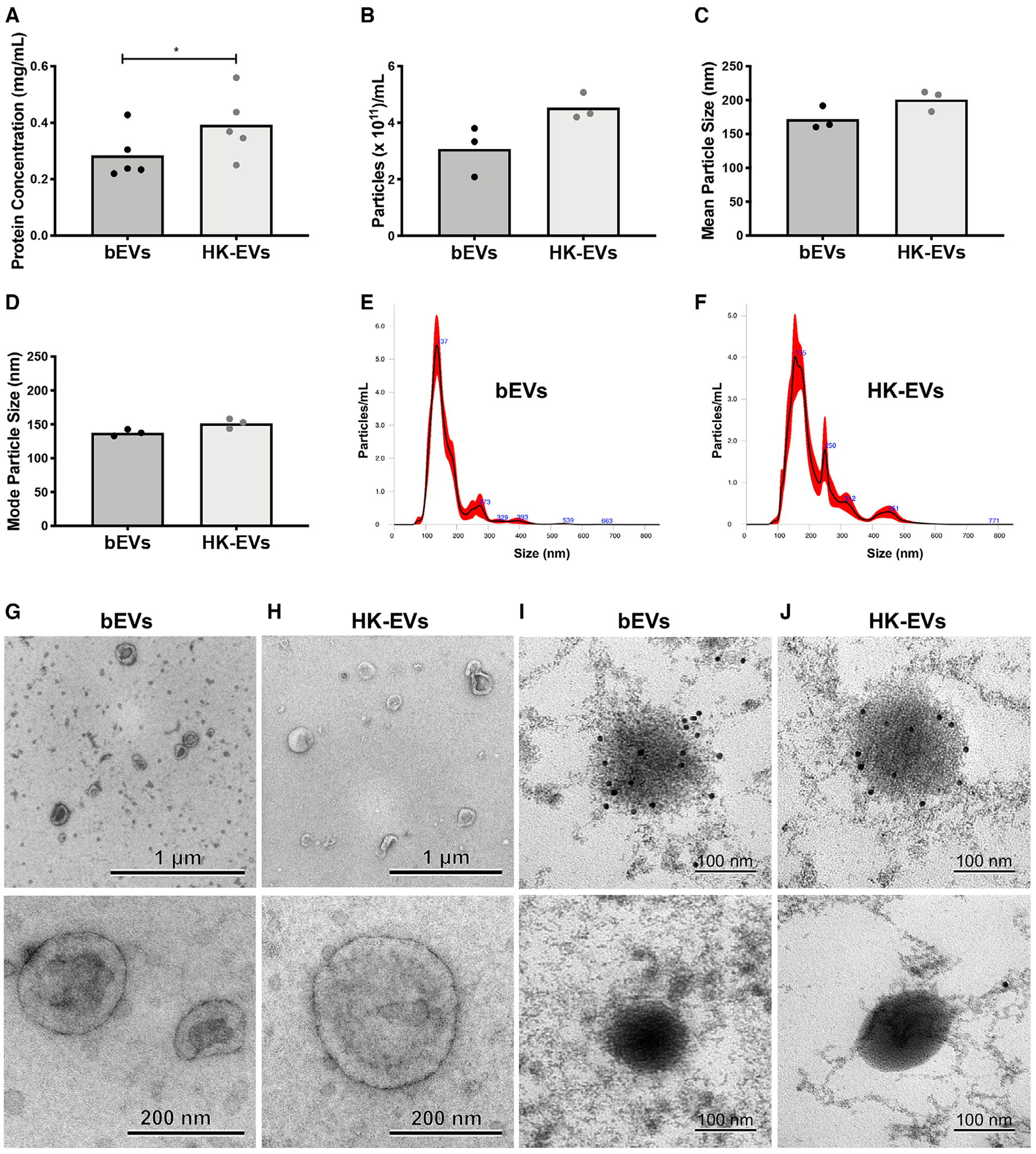
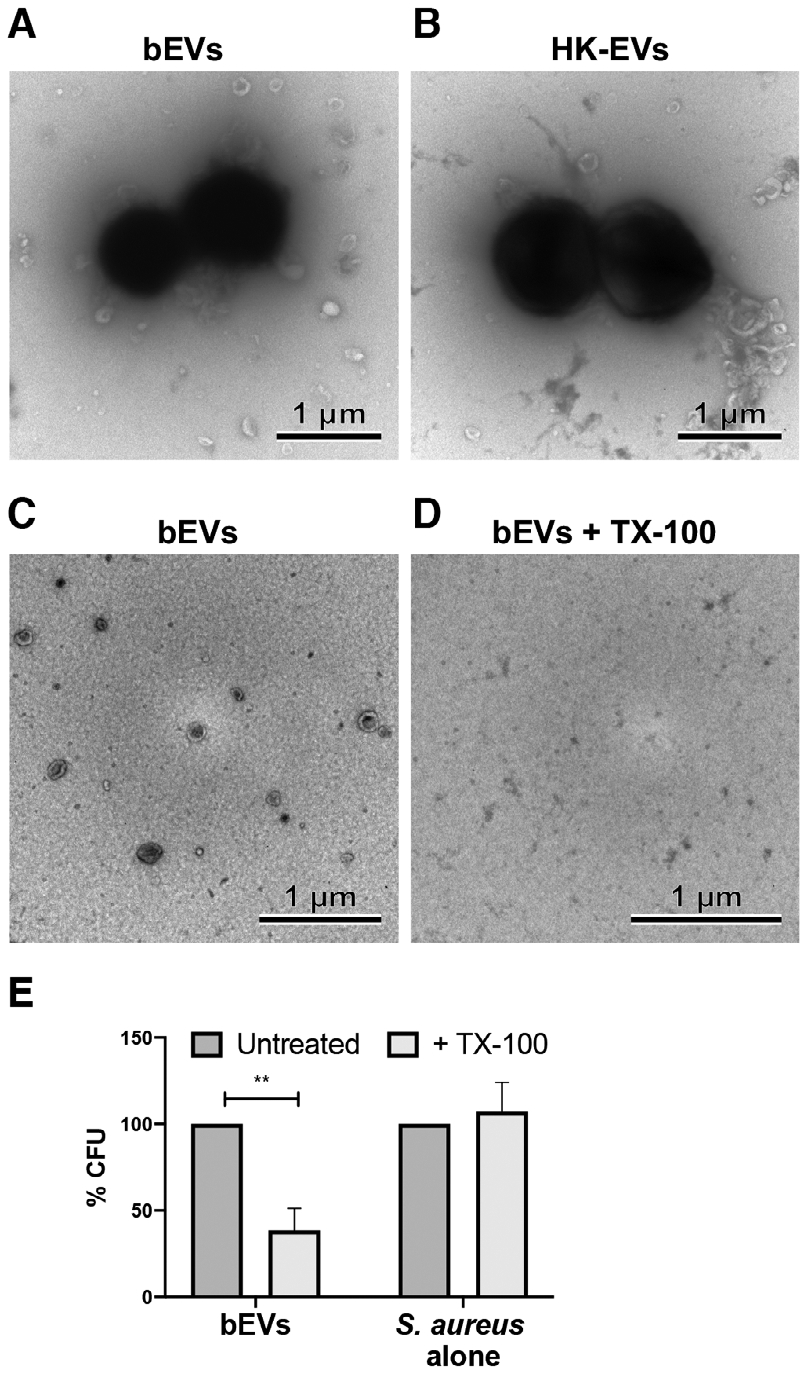
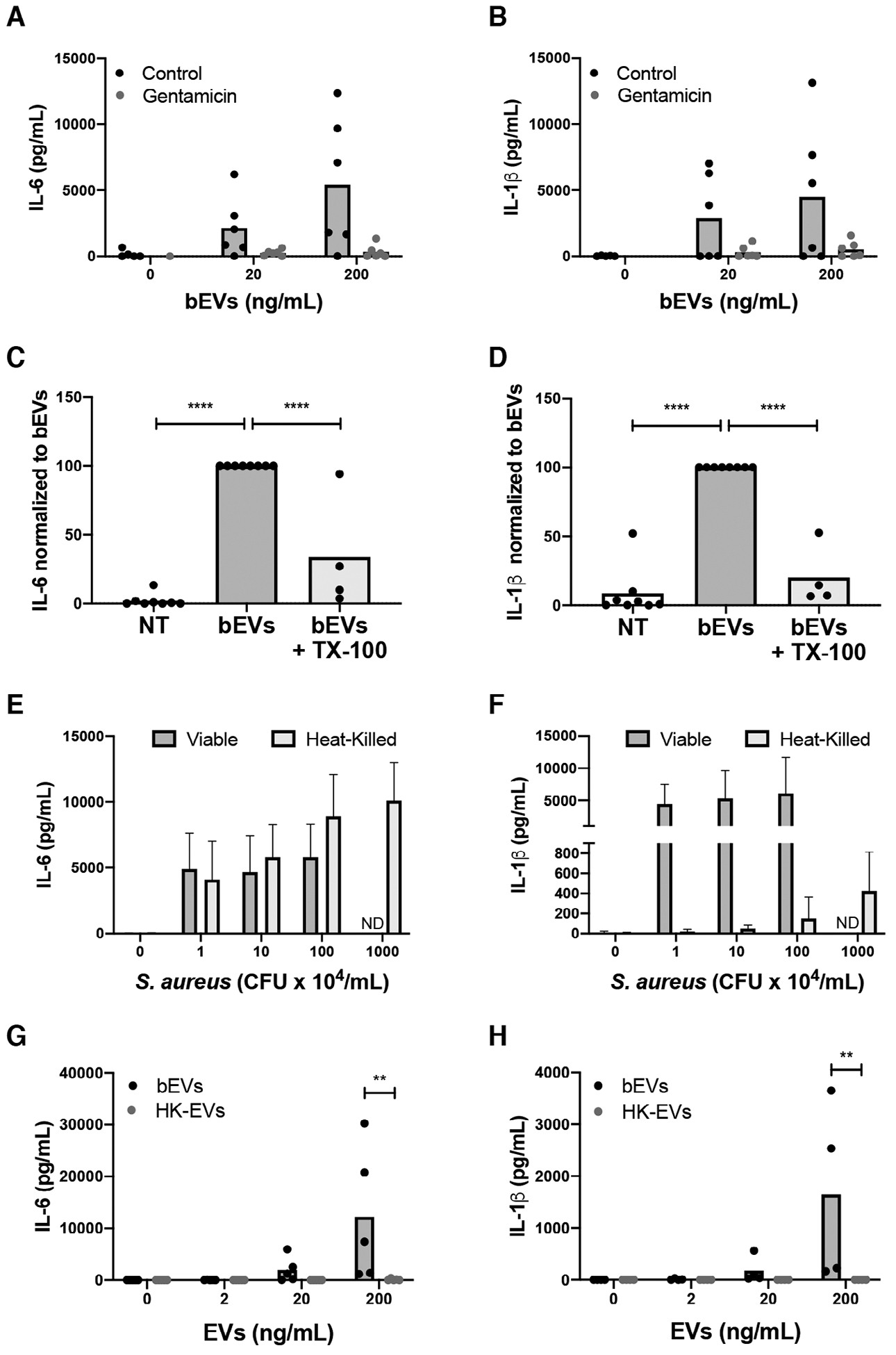
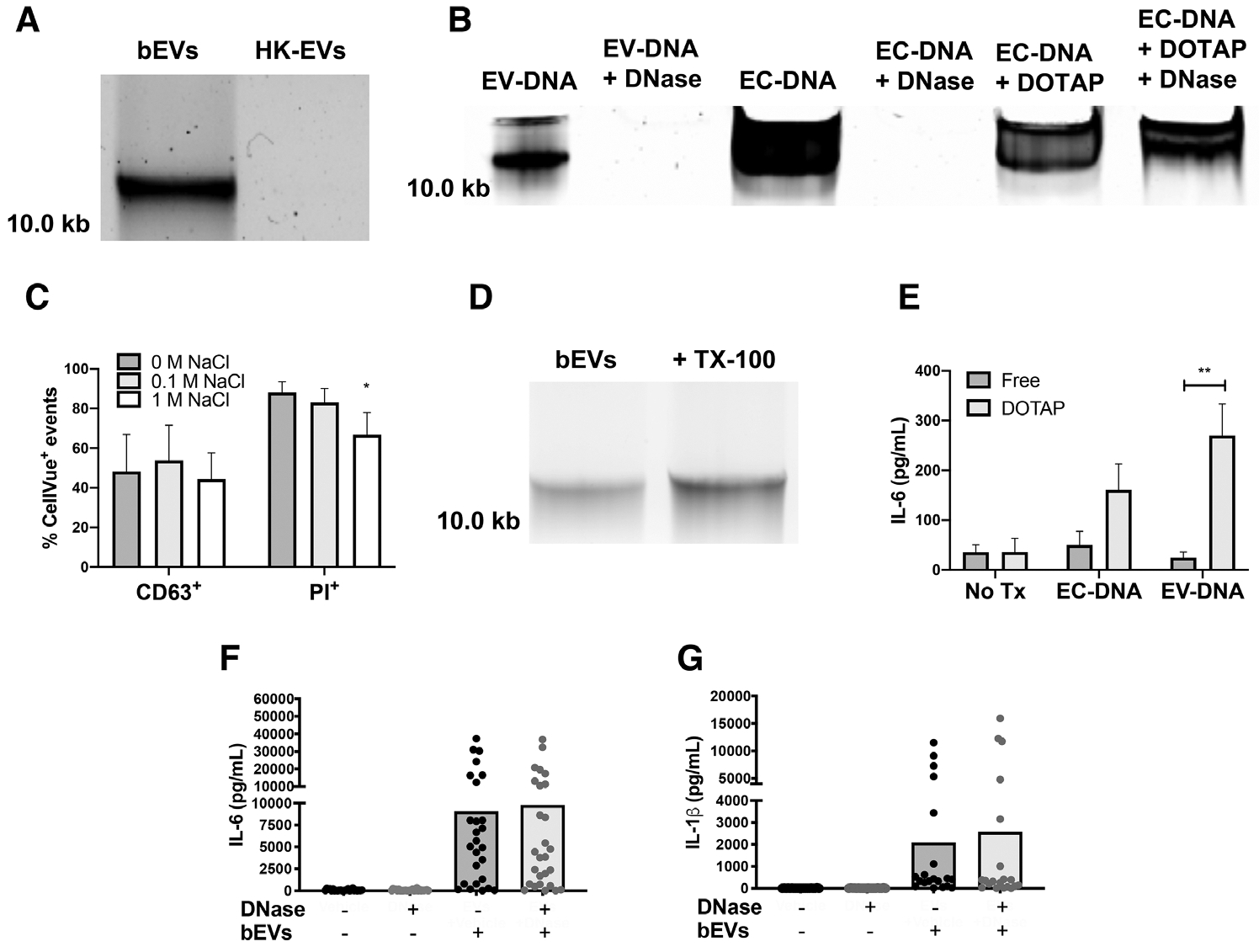
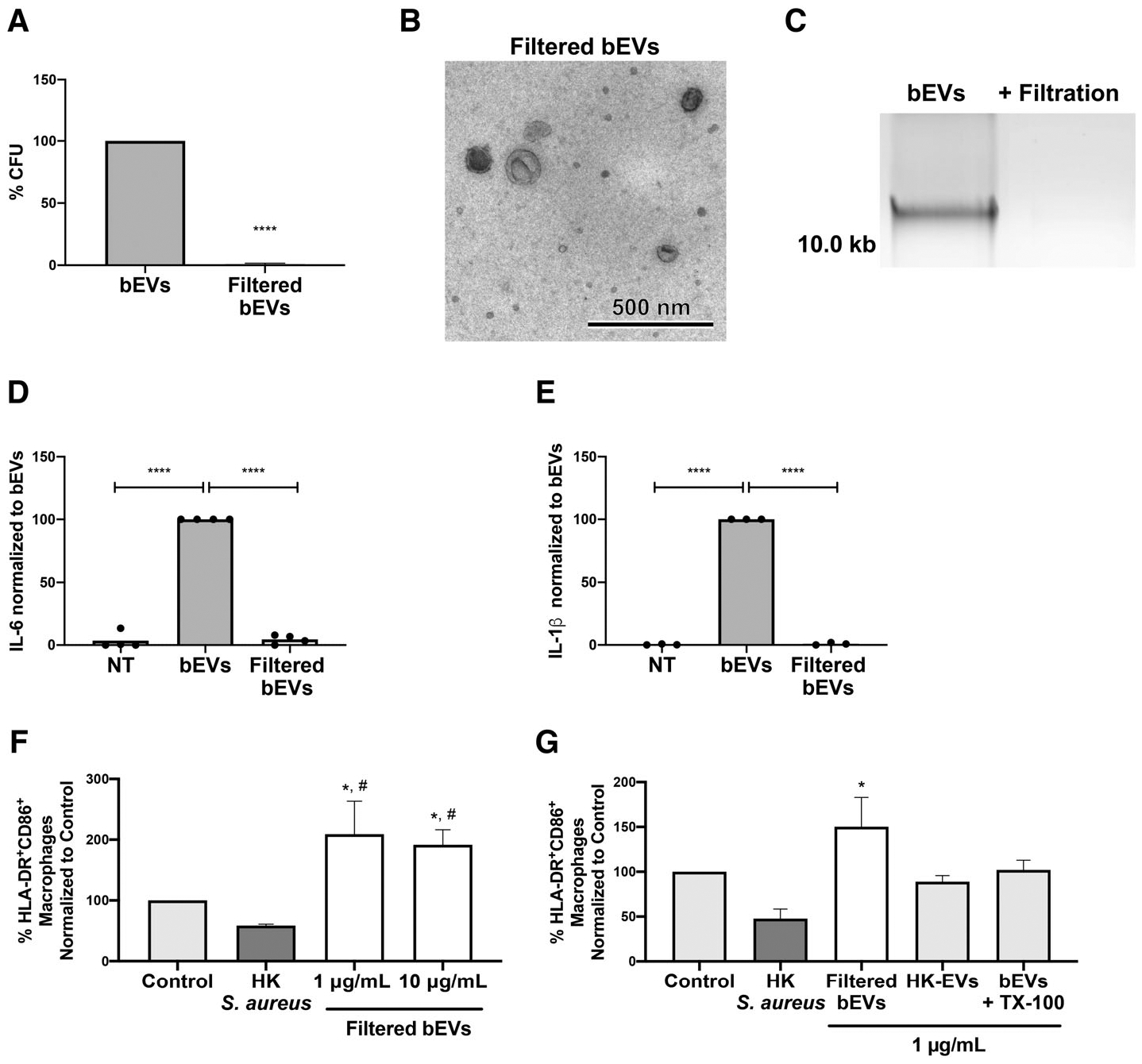
Staphylococcus aureus enhances neutrophil extracellular vesicle (EV) production. To investigate whether S. aureus viability influences EV biogenesis, EVs were isolated from human neutrophils incubated with viable bacteria (bEVs) or heat-killed bacteria (heat-killed EVs). Protein analysis, nanoparticle tracking and transmission electron microscopy showed comparable EV production between subsets, and both viable and nonviable bacteria were also detected in respective EV subsets. As anticipated, S. aureus, as well as bEVs with viable bacteria, were proinflammatory, and killing bacteria with gentamicin reduced cytokine production to baseline levels. Although heat-killed bacteria induced macrophage IL-6 production, heat-killed EVs did not. Additionally, we found that human and bacterial DNA associated with bEVs, but not heat-killed EVs, and that the DNA association could be partially decreased by disrupting electrostatic interactions. We investigated the potential for DNA isolated from EVs (EV-DNA) or EVs to cause inflammation. Although liposomal encapsulation of EV-DNA increased IL-6 production from baseline by 7.5-fold, treatment of bEVs with DNase I had no effect on IL-6 and IL-1β production, suggesting that the DNA did not contribute to the inflammatory response. Filtered EVs, which lacked DNA and associated bacteria, exhibited less proinflammatory activity relative to bEVs, and enhanced macrophage expression of CD86 and HLA-DR. Ultimately, we show that bEVs isolated by differential centrifugation co-purify with bacteria and DNA, and studying their concerted activity and relative contribution to immune response is important to the study of host-pathogen interactions.
Keywords: CD86 and HLA-DR; MRSA; ectosomes; exosomes; extracellular DNA; microparticles.
©2020 Society for Leukocyte Biology.
Conflict of interest statement
DISCLOSURES
The authors declare no conflicts of interest.
Figures
References
-
- Monaco M, Pimentel de Araujo F, Cruciani M, Coccia EM, Pantosti A. Worldwide epidemiology and antibiotic resistance of Staphylococcus aureus. Curr Top Microbiol Immunol. 2017;409:21–56. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
