

Systemic dysfunction and plasticity of the immune macroenvironment in cancer models
- PMID: 32451499
- PMCID: PMC7384250
- DOI: 10.1038/s41591-020-0892-6
Systemic dysfunction and plasticity of the immune macroenvironment in cancer models
Erratum in
-
Author Correction: Systemic dysfunction and plasticity of the immune macroenvironment in cancer models.Nat Med. 2024 May;30(5):1502. doi: 10.1038/s41591-024-02947-2. Nat Med. 2024. PMID: 38565647 No abstract available.
Abstract
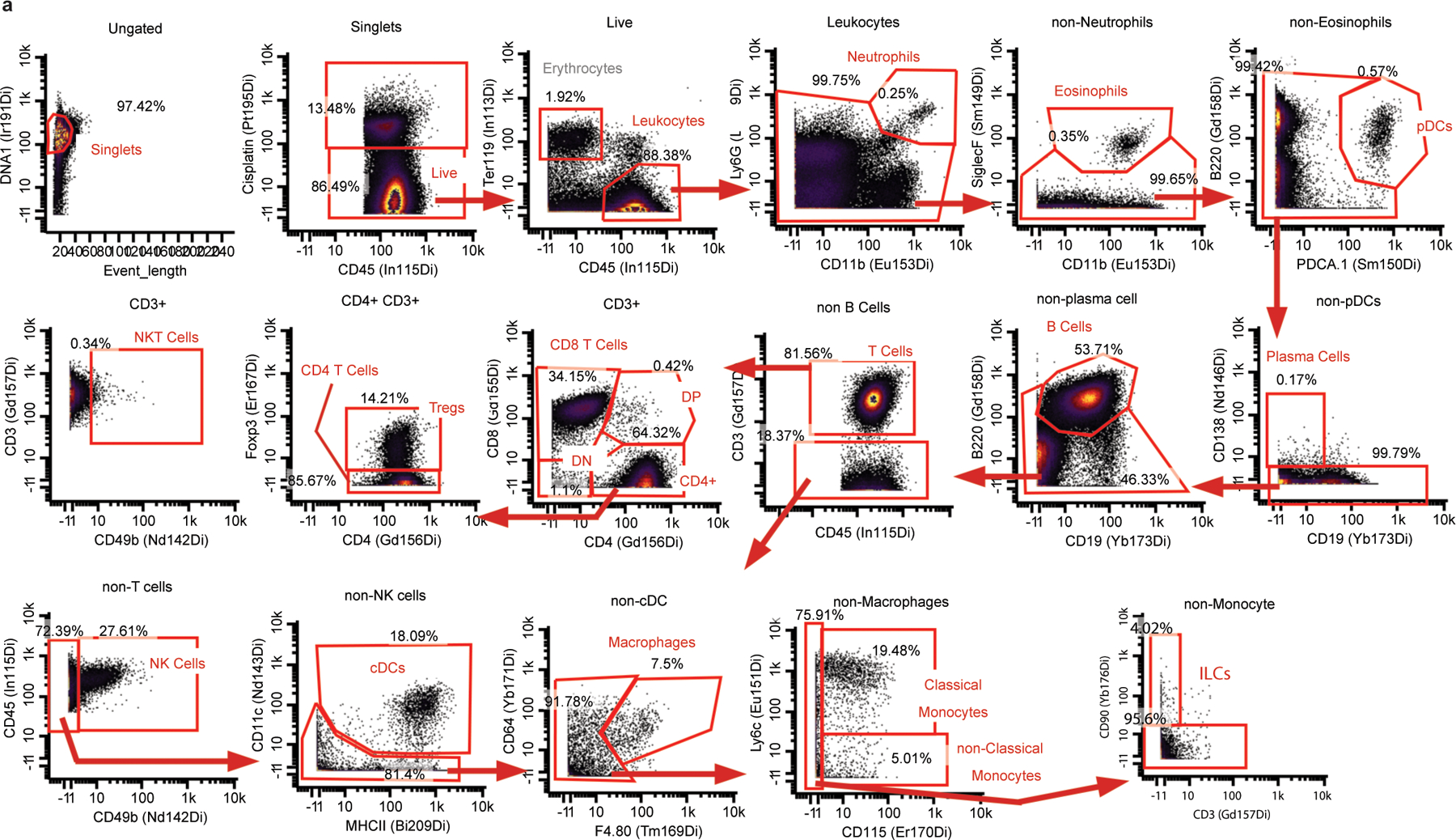
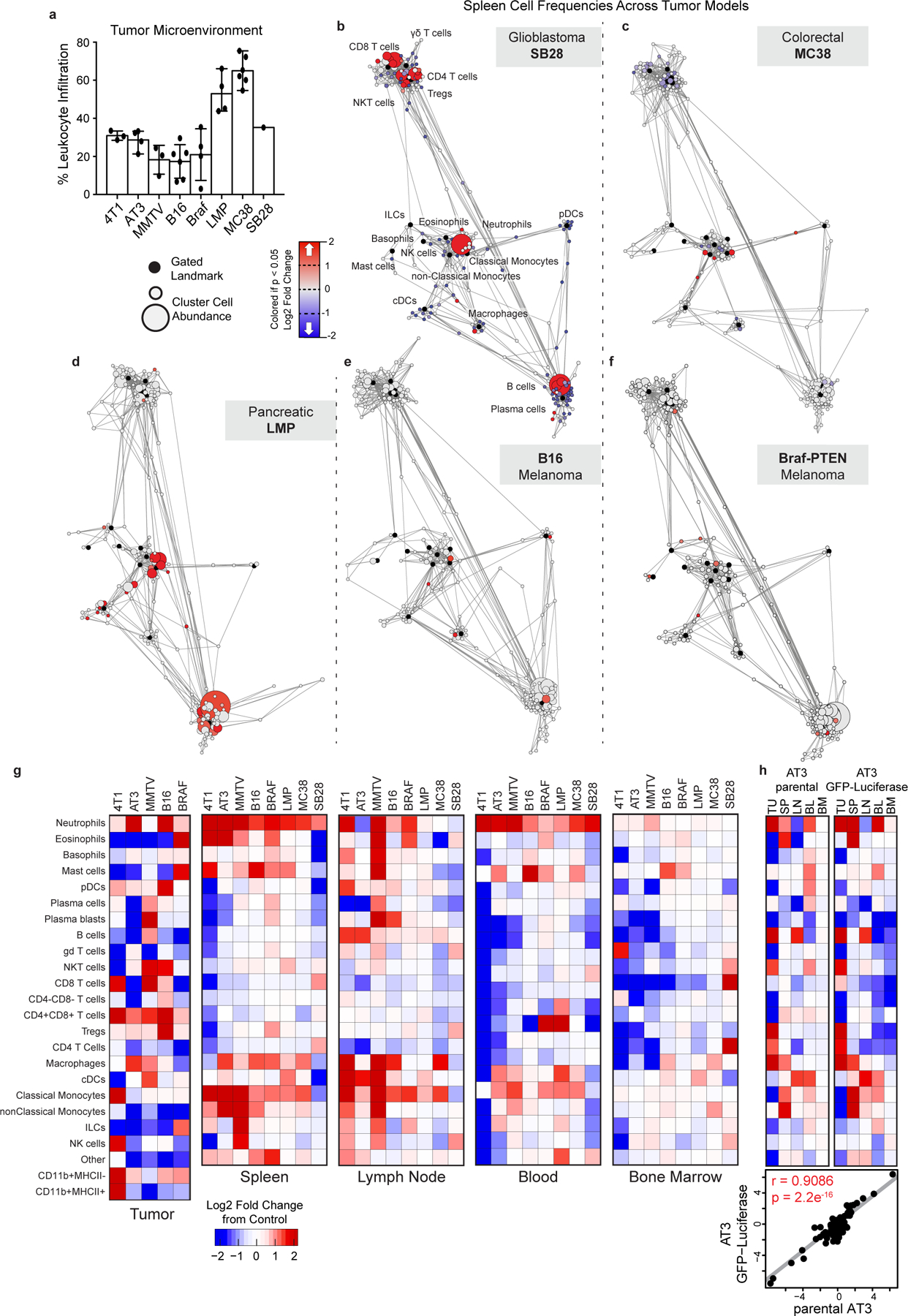
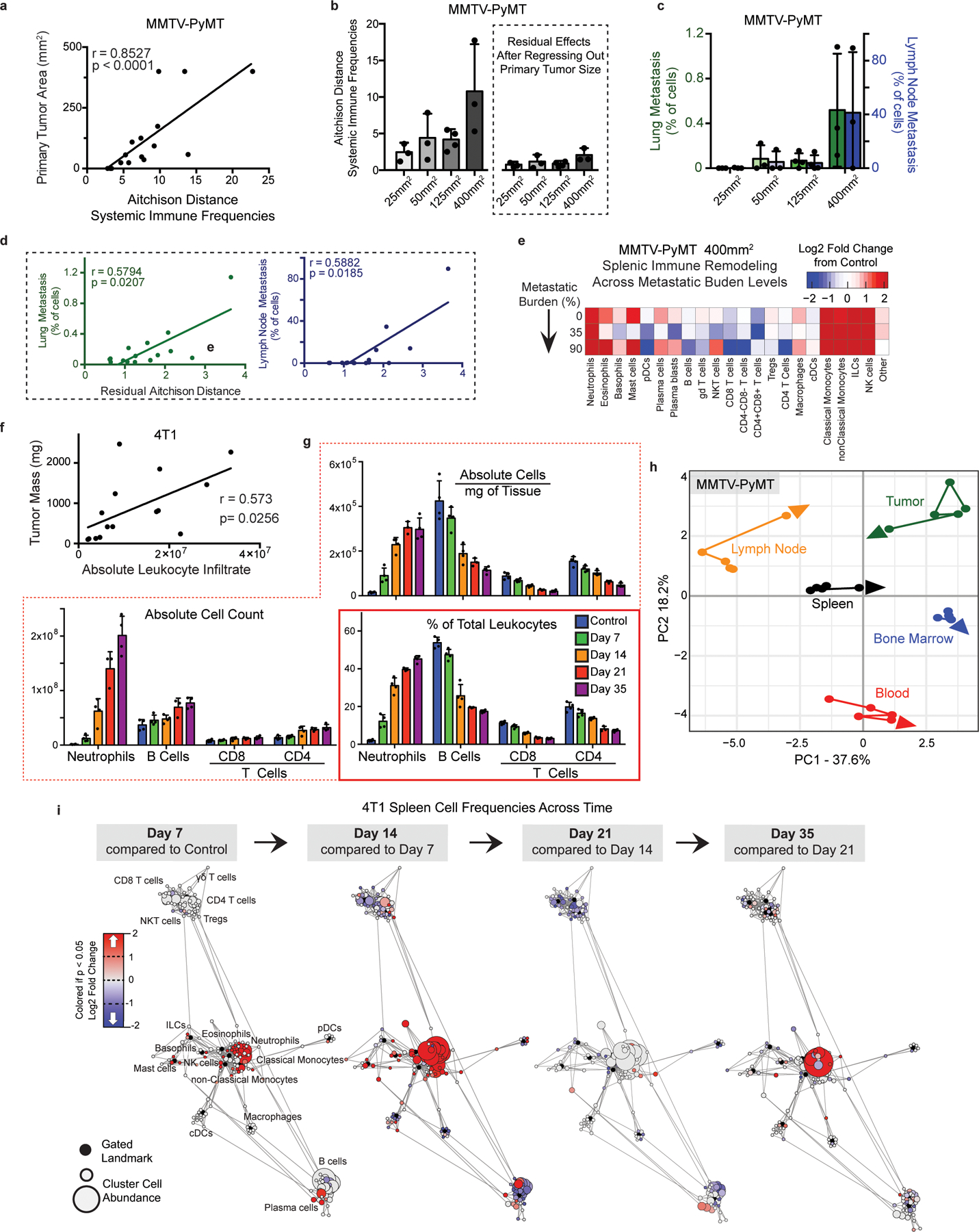
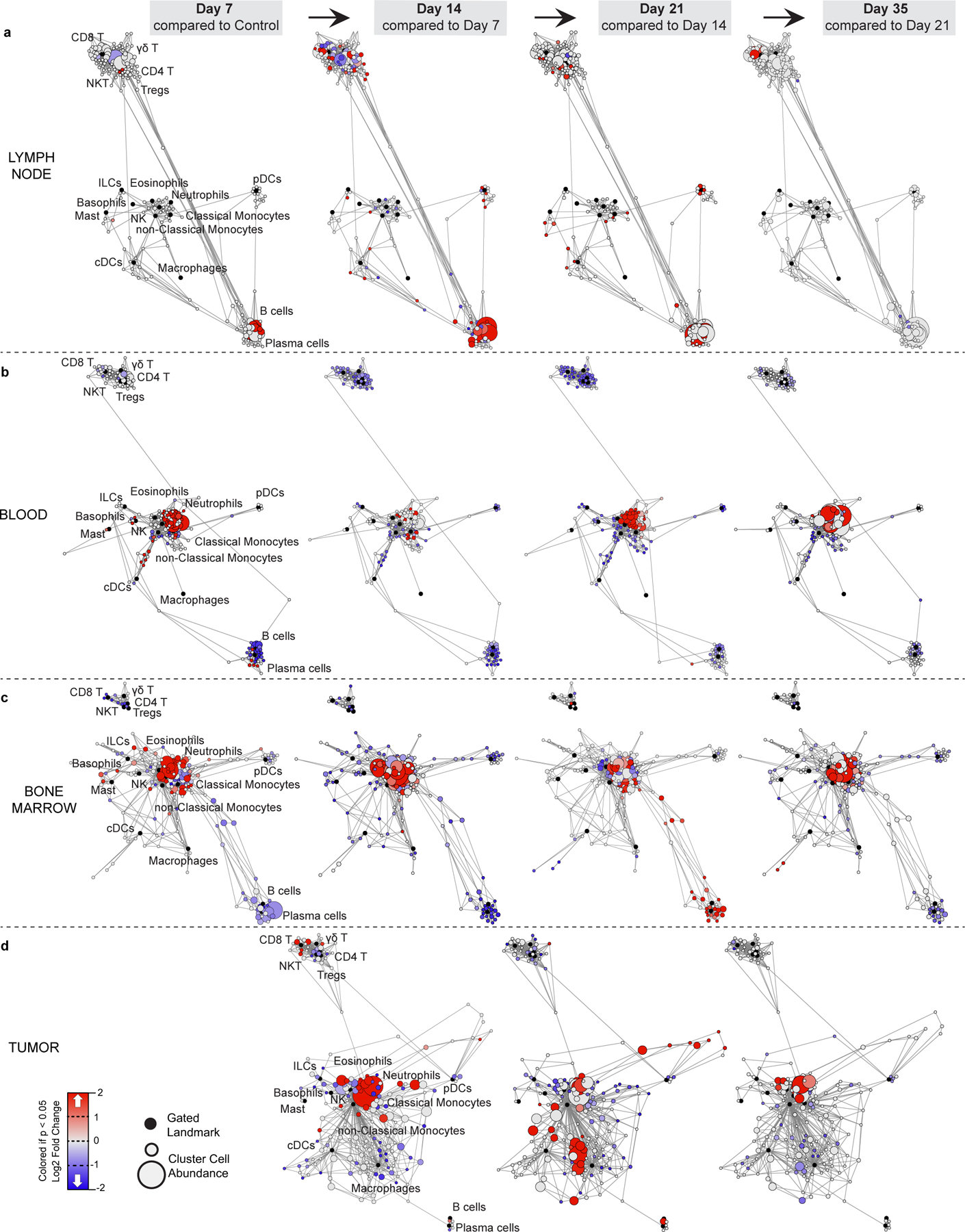
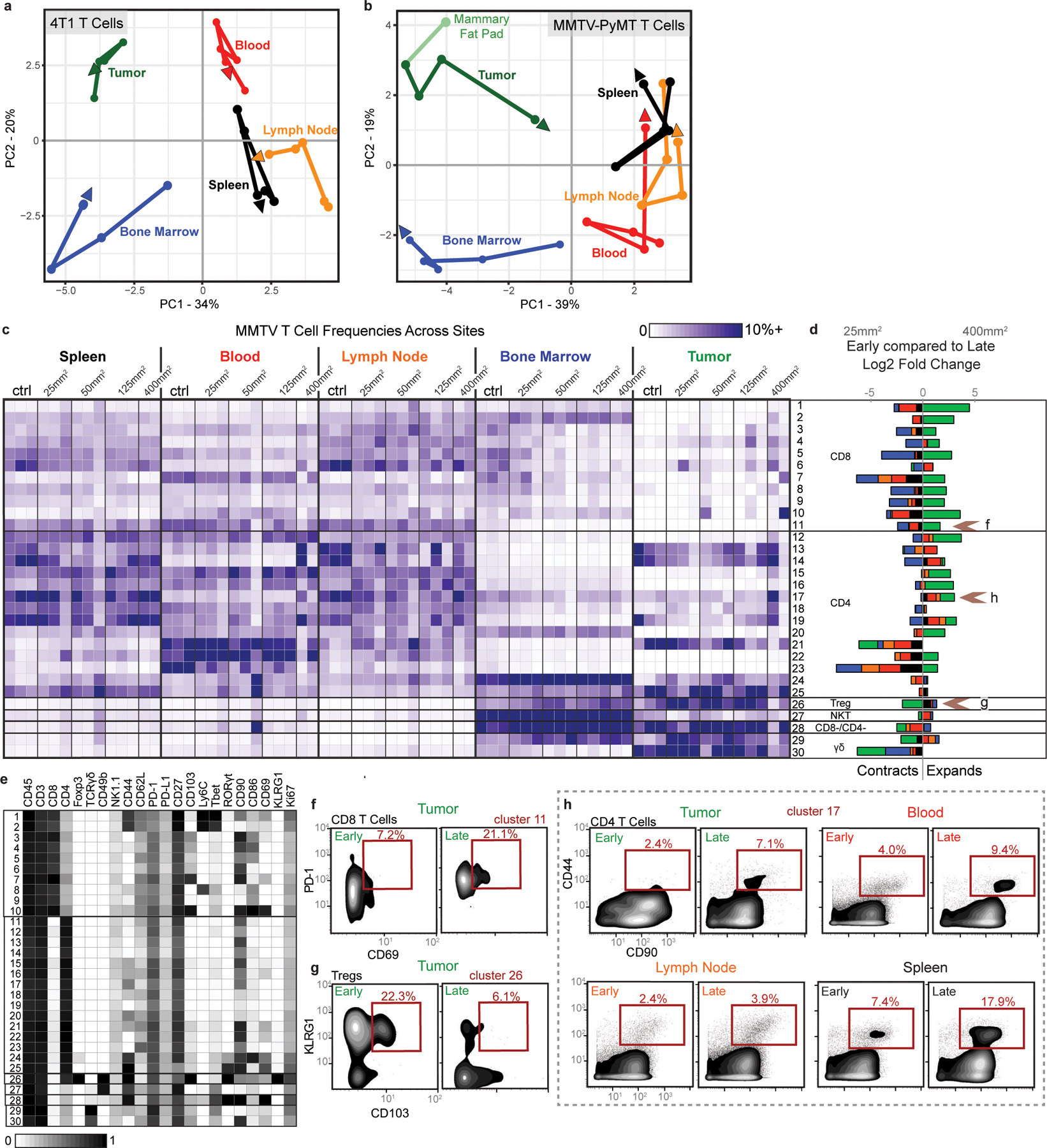
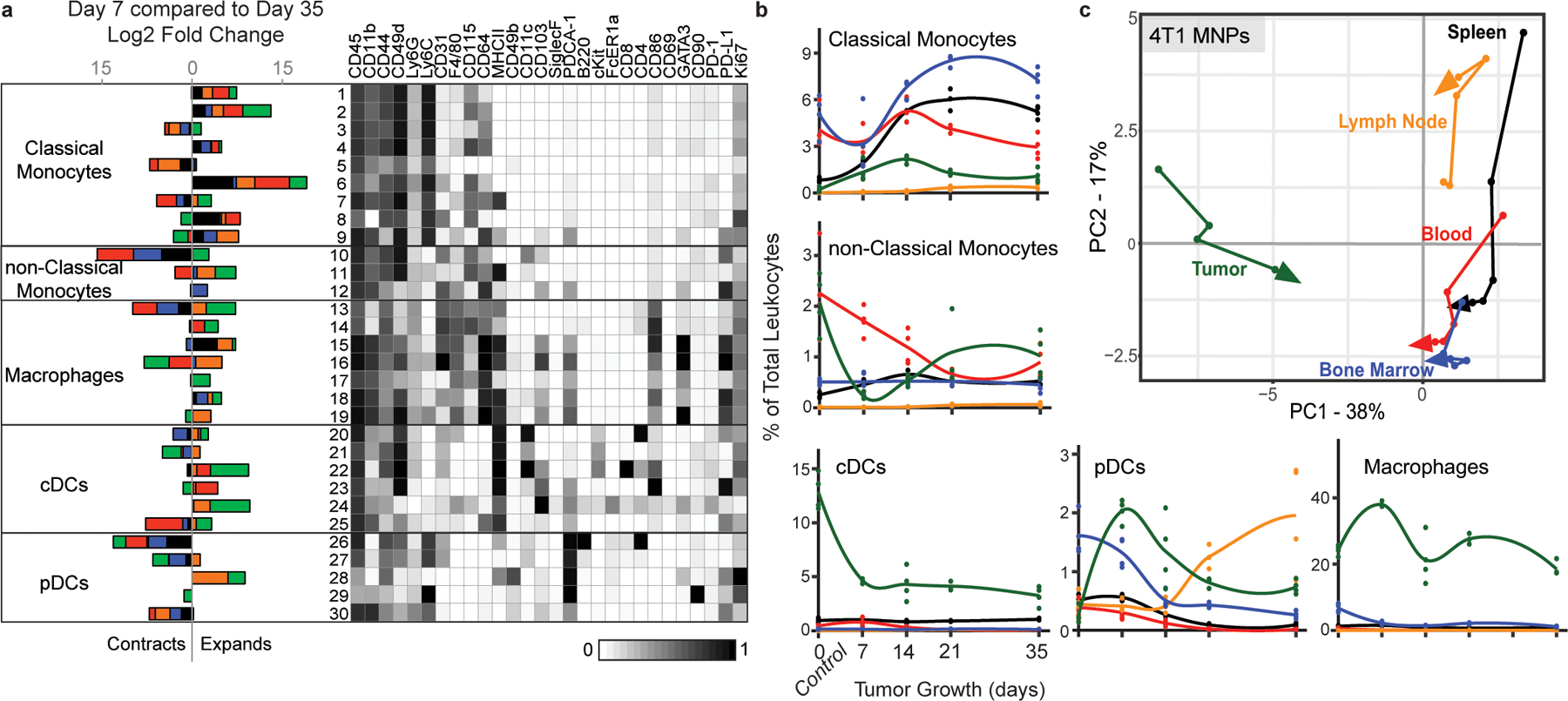
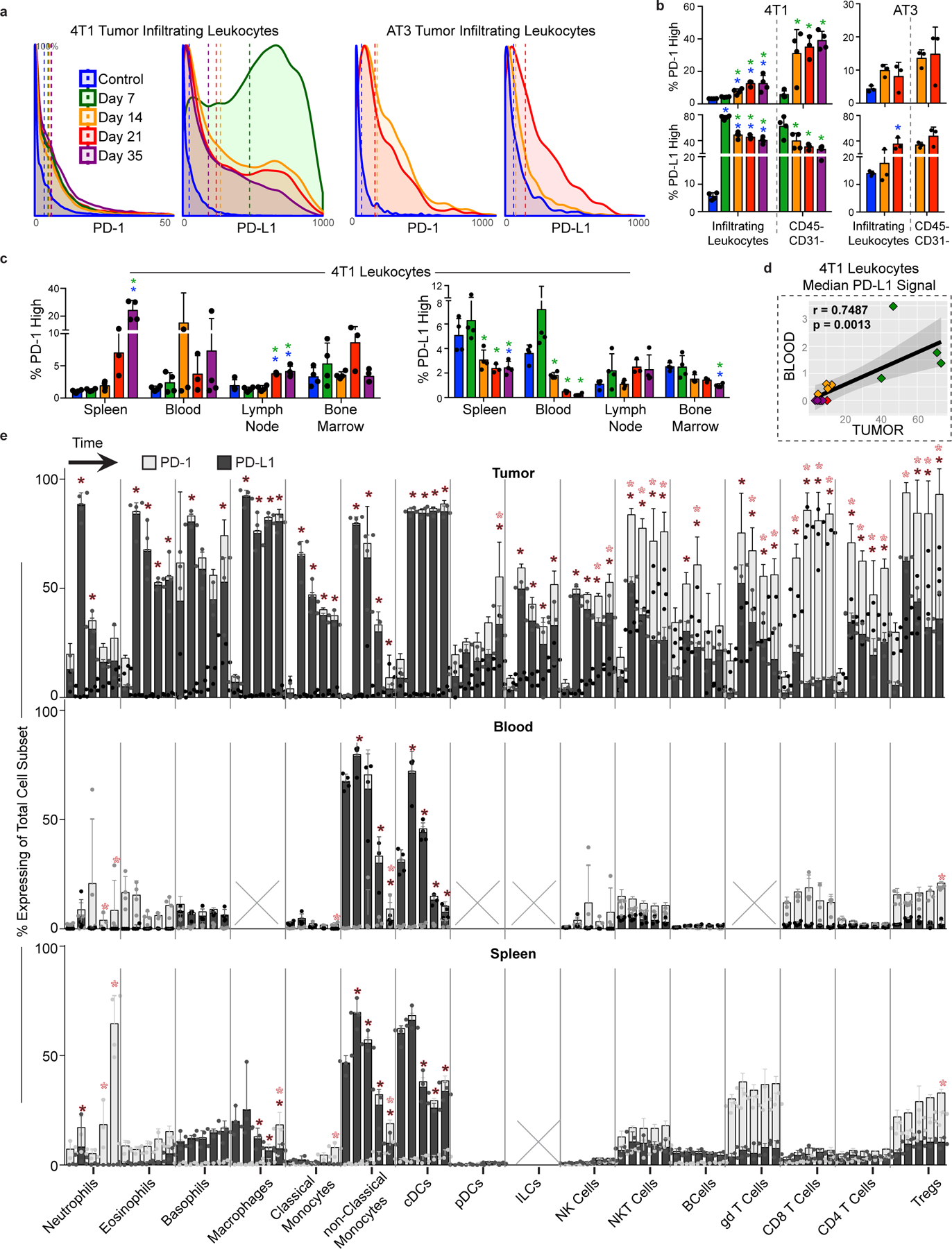
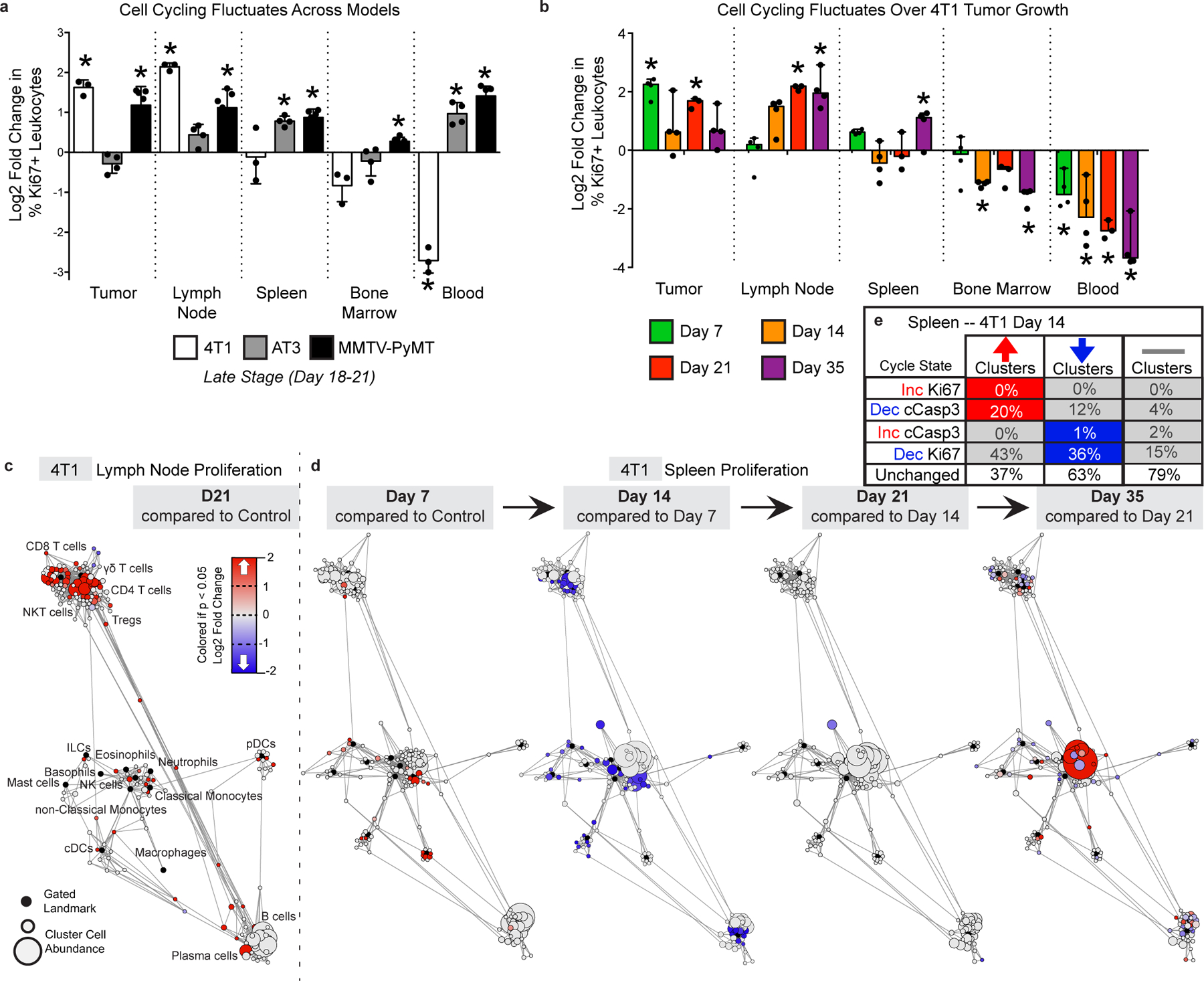
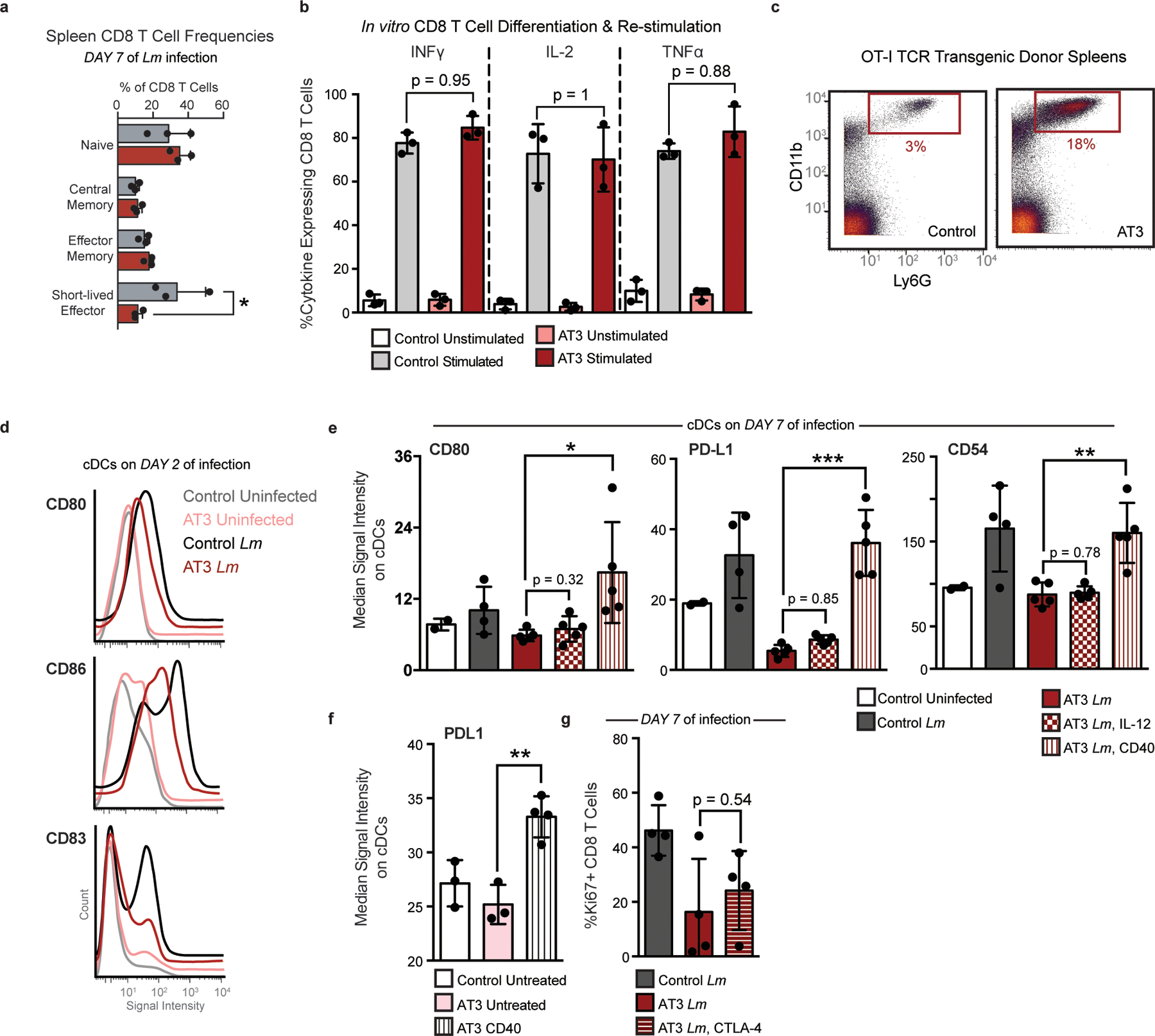
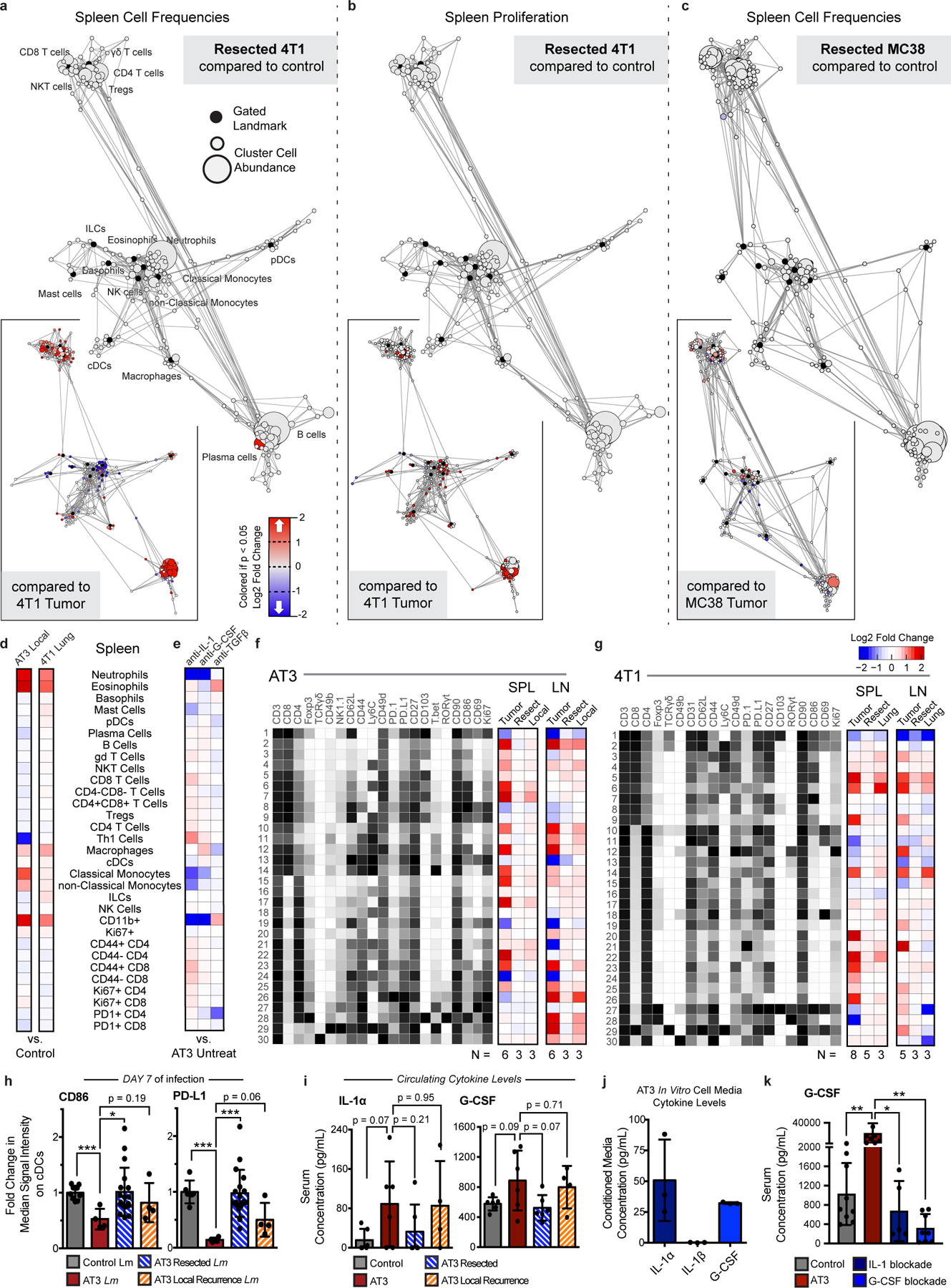
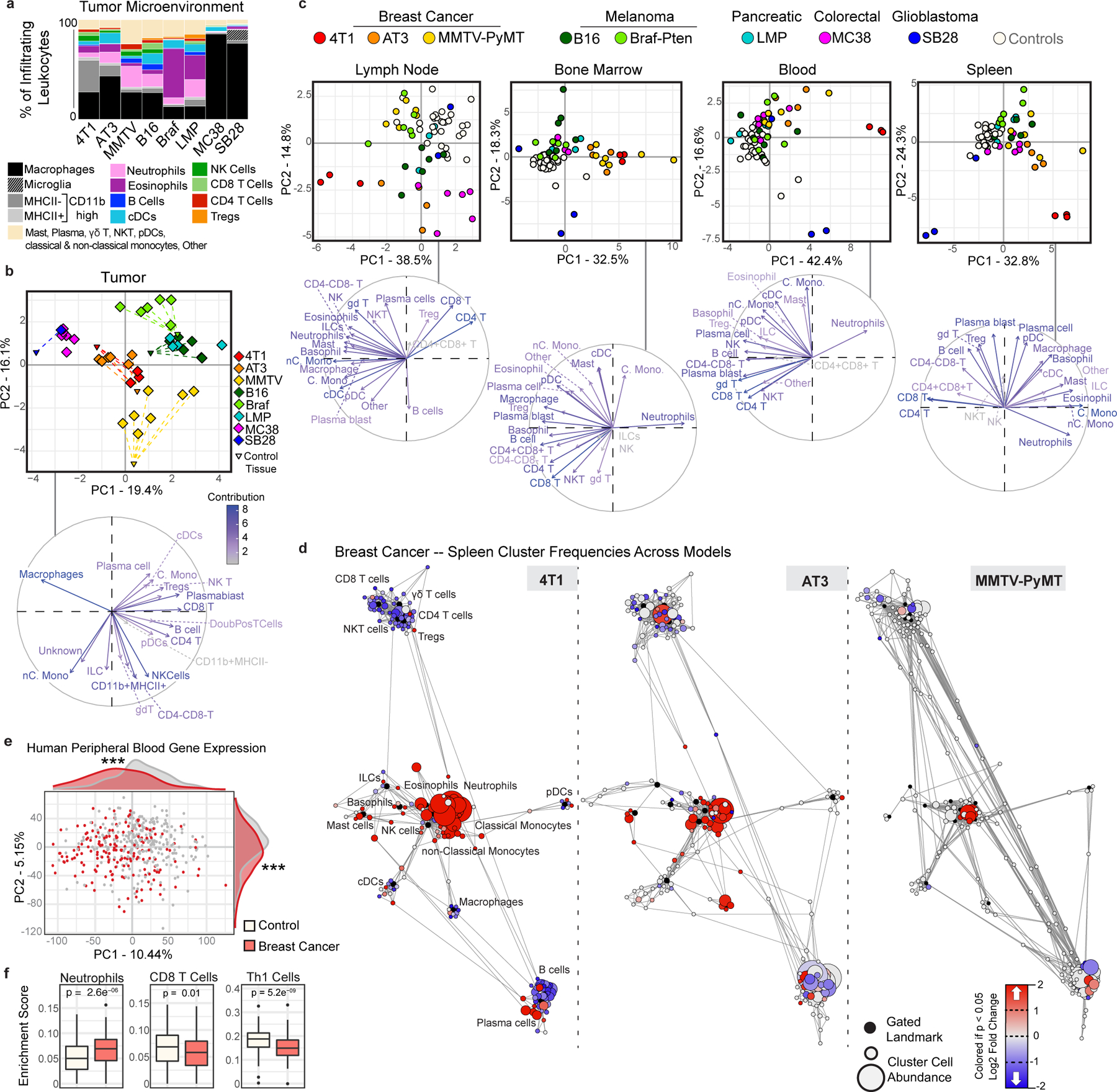
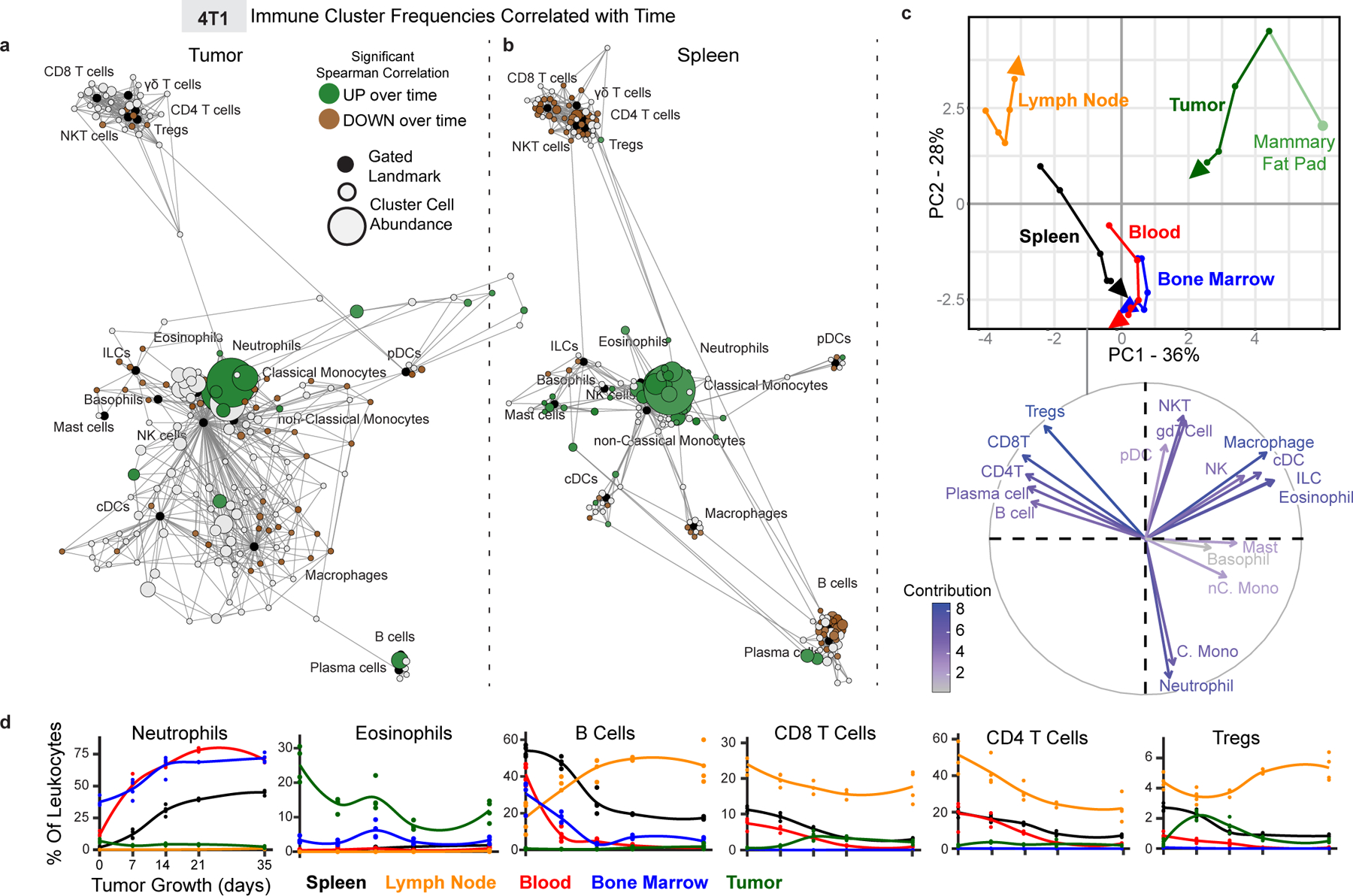
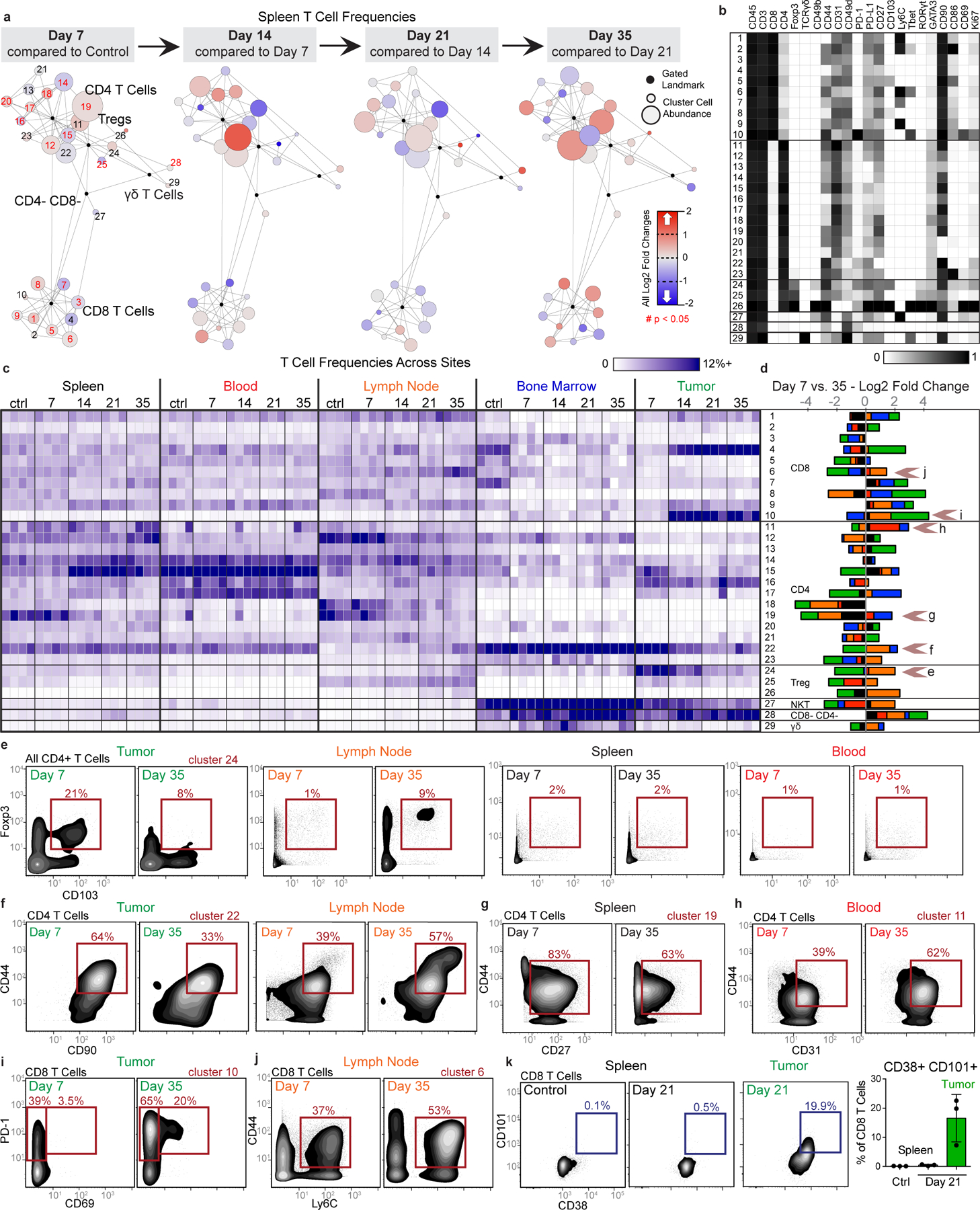
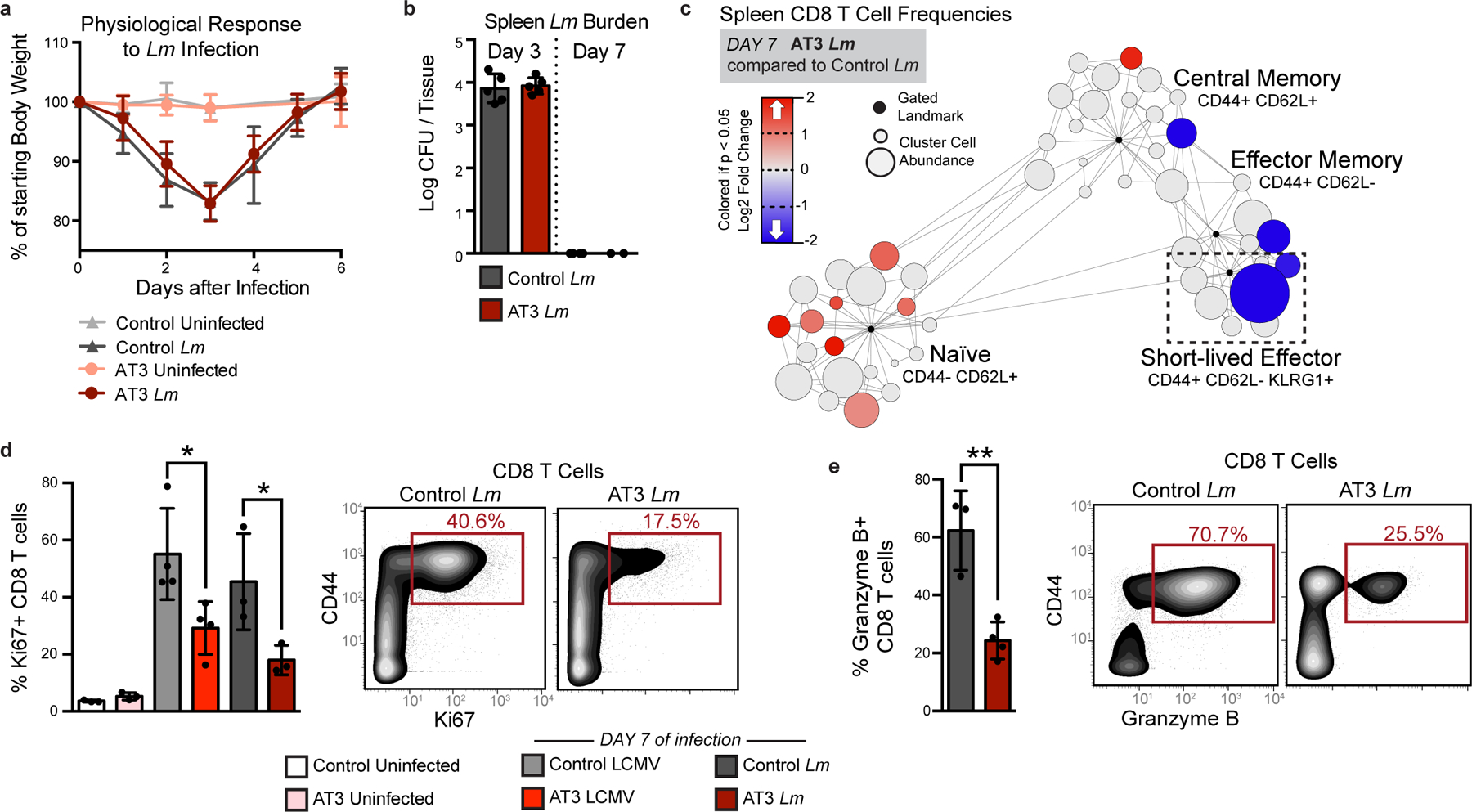
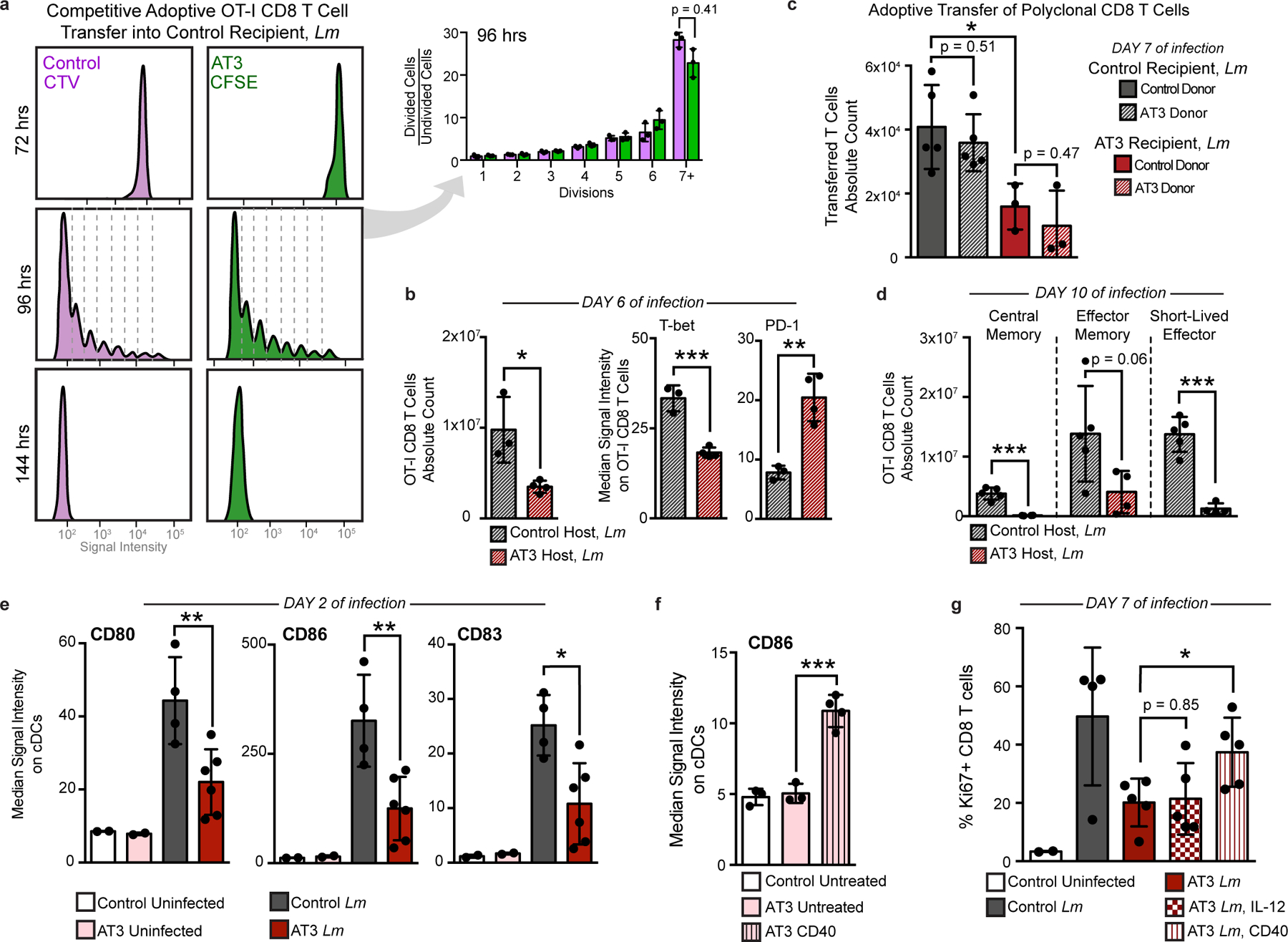
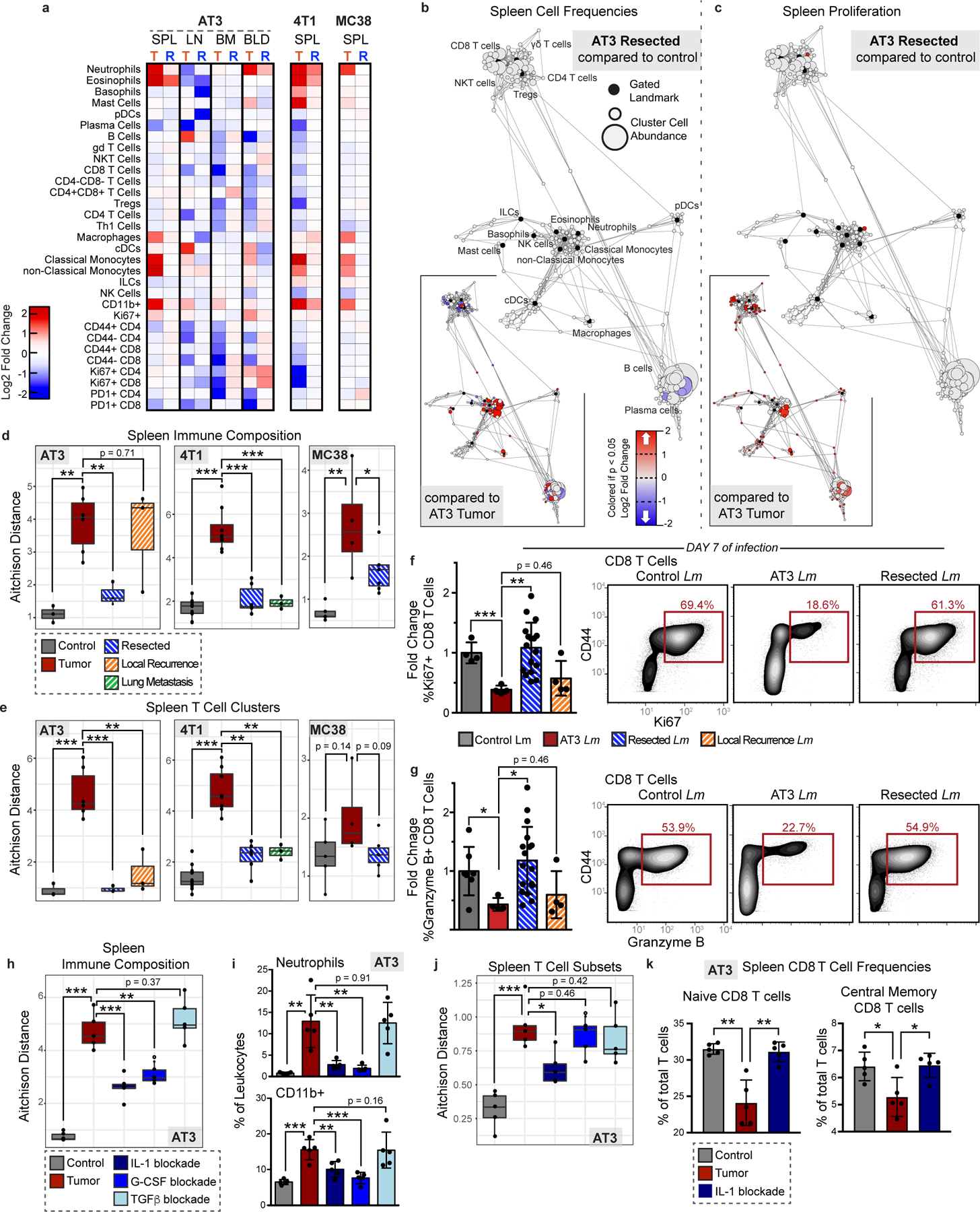
Understanding of the factors governing immune responses in cancer remains incomplete, limiting patient benefit. In this study, we used mass cytometry to define the systemic immune landscape in response to tumor development across five tissues in eight mouse tumor models. Systemic immunity was dramatically altered across models and time, with consistent findings in the peripheral blood of patients with breast cancer. Changes in peripheral tissues differed from those in the tumor microenvironment. Mice with tumor-experienced immune systems mounted dampened responses to orthogonal challenges, including reduced T cell activation during viral or bacterial infection. Antigen-presenting cells (APCs) mounted weaker responses in this context, whereas promoting APC activation rescued T cell activity. Systemic immune changes were reversed with surgical tumor resection, and many were prevented by interleukin-1 or granulocyte colony-stimulating factor blockade, revealing remarkable plasticity in the systemic immune state. These results demonstrate that tumor development dynamically reshapes the composition and function of the immune macroenvironment.
Figures
Comment in
-
Tumours disrupt the immune scenery.Nat Rev Cancer. 2020 Aug;20(8):415. doi: 10.1038/s41568-020-0286-6. Nat Rev Cancer. 2020. PMID: 32620876 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
