

Pretreatment of Anthocyanin from the Fruit of Vitis coignetiae Pulliat Acts as a Potent Inhibitor of TNF-α Effect by Inhibiting NF-κB-Regulated Genes in Human Breast Cancer Cells
- PMID: 32455624
- PMCID: PMC7287973
- DOI: 10.3390/molecules25102396
Pretreatment of Anthocyanin from the Fruit of Vitis coignetiae Pulliat Acts as a Potent Inhibitor of TNF-α Effect by Inhibiting NF-κB-Regulated Genes in Human Breast Cancer Cells
Abstract
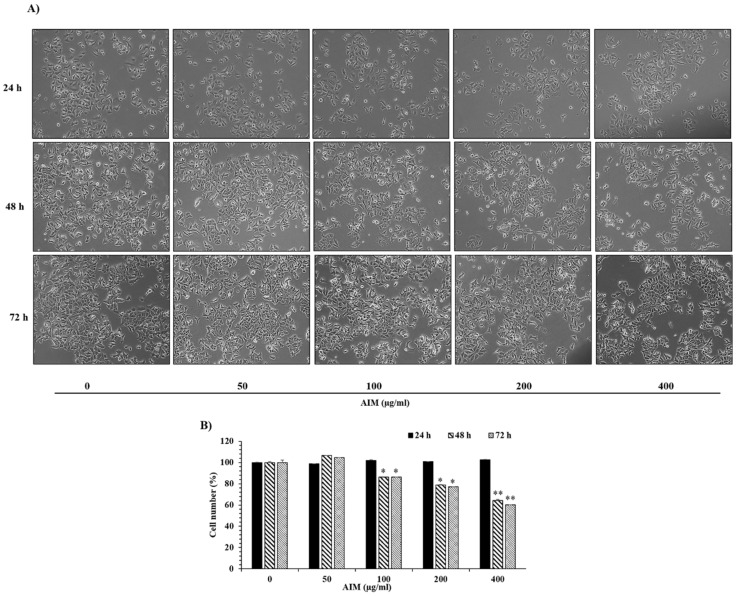
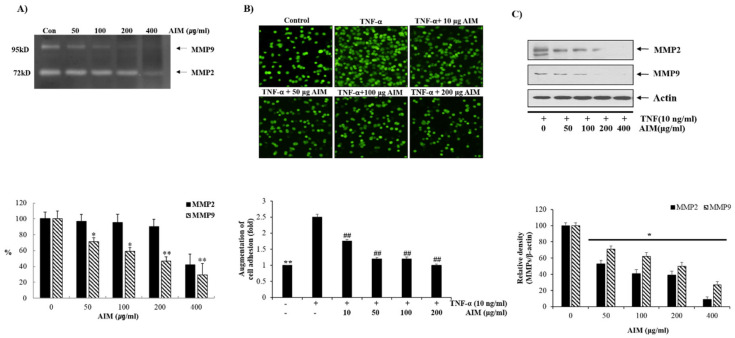
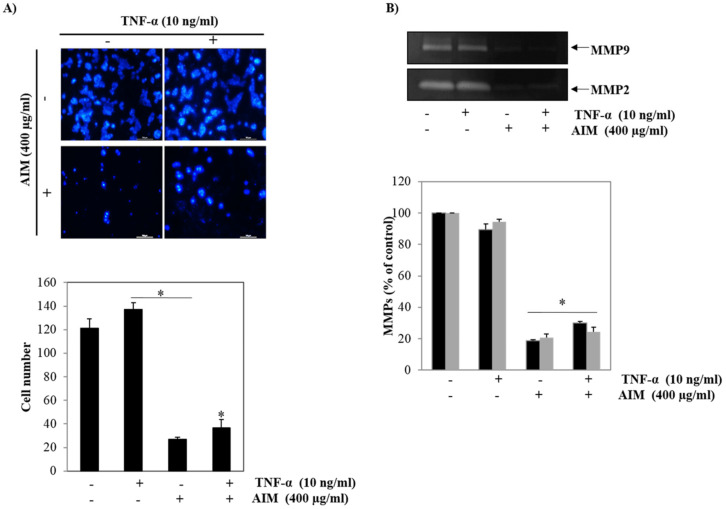
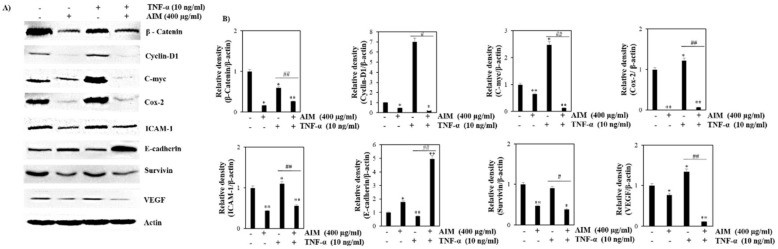
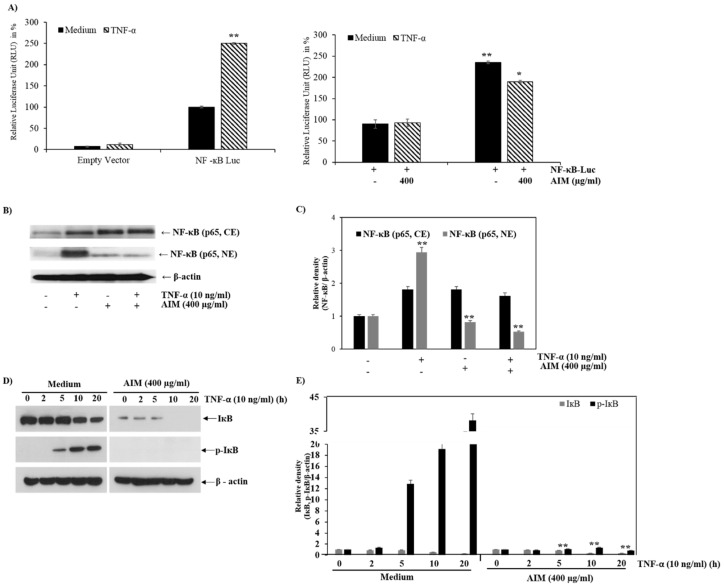
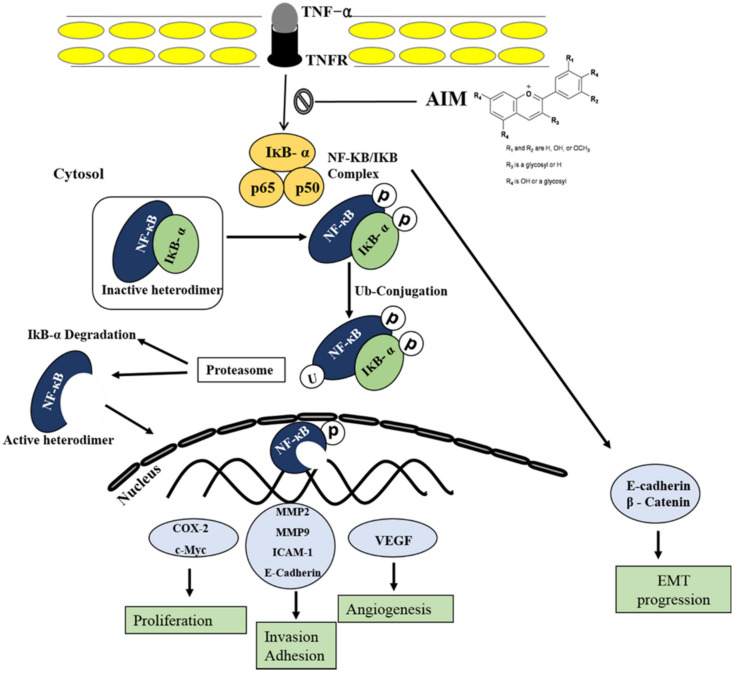
Vitis coignetiae Pulliat (Meoru in Korea) has been used in Korean folk medicine for the treatment of inflammatory diseases and cancers. Evidence suggests that NF-κB activation is mainly involved in cancer cell proliferation, invasion, angiogenesis, and metastasis. TNF-α also enhances the inflammatory process in tumor development. Recently, flavonoids from plants have been reported to have inhibitory effects on NF-κB activities. We investigated the effects of anthocyanins extracted from the fruits of Vitis coignetiae Pulliat (AIM, anthocyanins isolated from Meoru (AIM)) on TNF-α-induced NF-κB activities in MCF-7 human breast cancer cells and the molecules involved in AIM-induced anti-cancer effects, especially on cancer metastasis. We performed cell viability assay, gelatin zymography, invasion assay, and western blot analysis to unravel the anti-NF-κB activity of AIMs on MCF-7 cells. AIM suppressed the TNF-α effects on the NF-κB-regulated proteins involved in cancer cell proliferation (COX-2, C-myc), invasion, and angiogenesis (MMP-2, MMP9, ICAM-1, and VEGF). AIM also increased the expression of E-cadherin, which is one of the hallmarks of the epithelial-mesenchymal transition (EMT) process. In conclusion, this study demonstrates that the anthocyanins isolated from the fruits of Vitis coignetiae Pulliat acts as an inhibitor of TNF-α induced NF-κB activation, and subsequent downstream molecules involved in cancer proliferation, invasion, adhesion, angiogenesis, and thus have anti-metastatic activities in MCF-7 breast cancer cells.
Keywords: NF-κB; Vitis coignetiae Pulliat; anthocyanins; breast cancer.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- Comsa S., Cimpean A.M., Raica M. The story of MCF-7 breast cancer cell line: 40 years of experience in research. Anticancer Res. 2015;35:3147–3154. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
