

Plant Genome Editing and the Relevance of Off-Target Changes
- PMID: 32457089
- PMCID: PMC7401131
- DOI: 10.1104/pp.19.01194
Plant Genome Editing and the Relevance of Off-Target Changes
Abstract
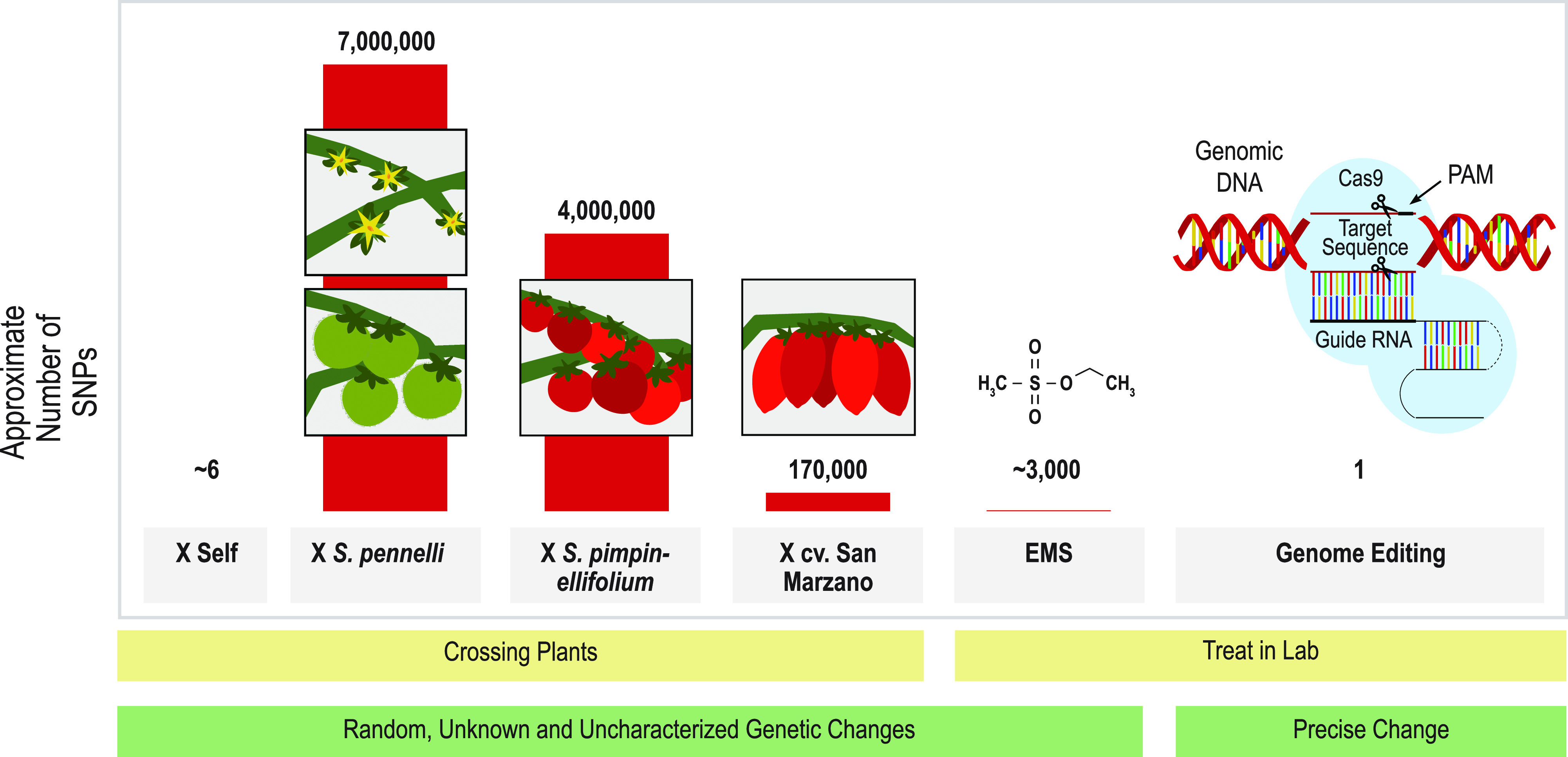
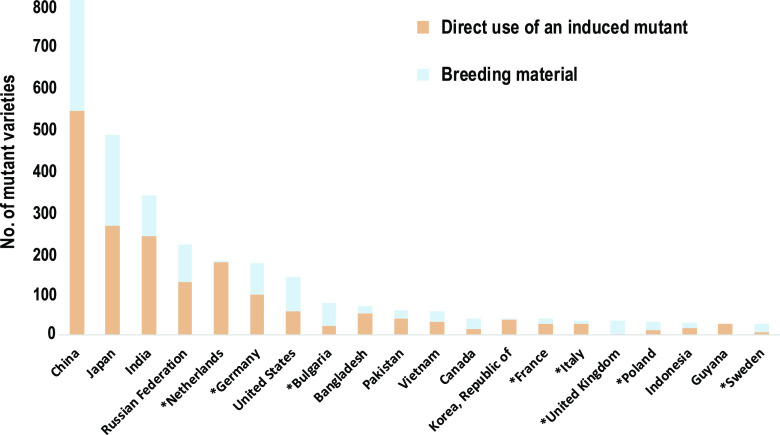
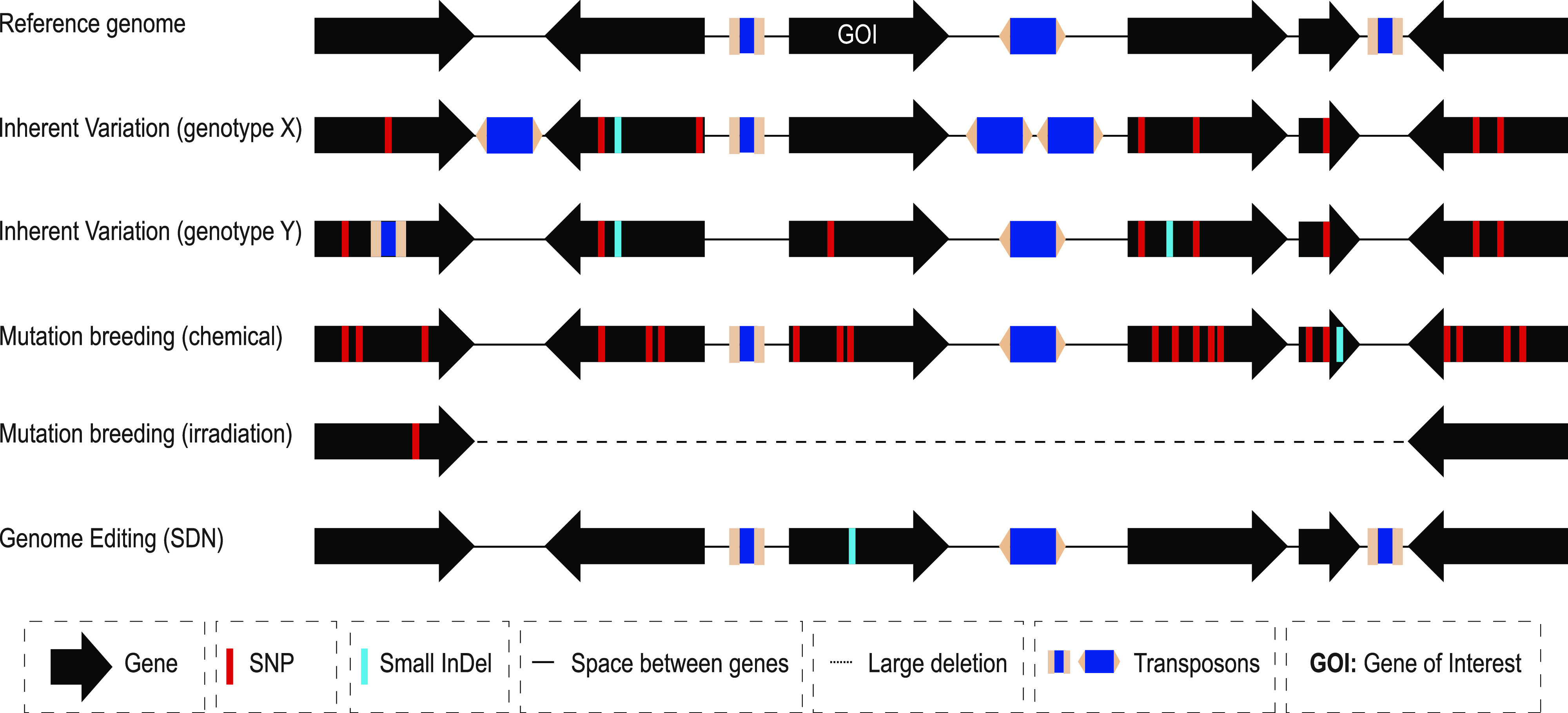
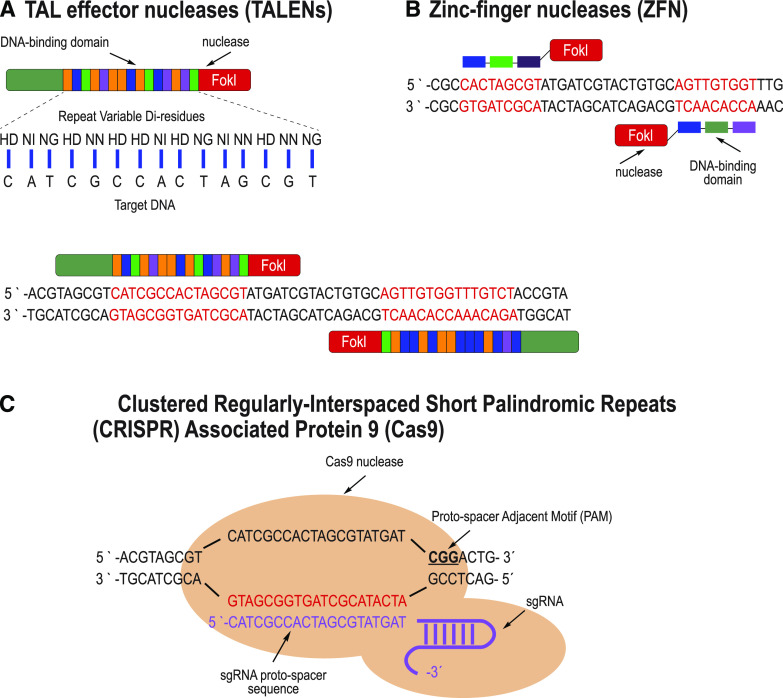
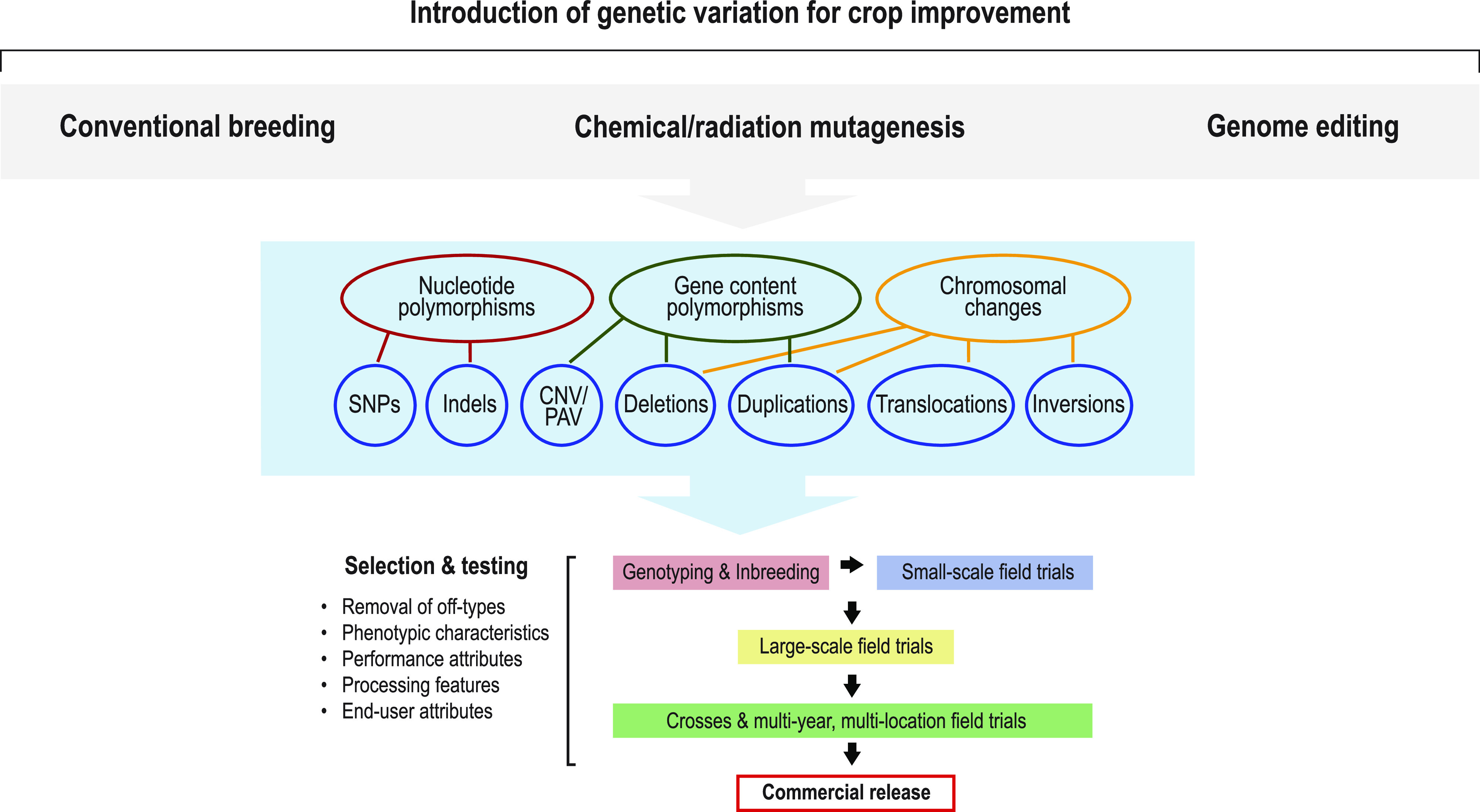
Site-directed nucleases (SDNs) used for targeted genome editing are powerful new tools to introduce precise genetic changes into plants. Like traditional approaches, such as conventional crossing and induced mutagenesis, genome editing aims to improve crop yield and nutrition. Next-generation sequencing studies demonstrate that across their genomes, populations of crop species typically carry millions of single nucleotide polymorphisms and many copy number and structural variants. Spontaneous mutations occur at rates of ∼10-8 to 10-9 per site per generation, while variation induced by chemical treatment or ionizing radiation results in higher mutation rates. In the context of SDNs, an off-target change or edit is an unintended, nonspecific mutation occurring at a site with sequence similarity to the targeted edit region. SDN-mediated off-target changes can contribute to a small number of additional genetic variants compared to those that occur naturally in breeding populations or are introduced by induced-mutagenesis methods. Recent studies show that using computational algorithms to design genome editing reagents can mitigate off-target edits in plants. Finally, crops are subject to strong selection to eliminate off-type plants through well-established multigenerational breeding, selection, and commercial variety development practices. Within this context, off-target edits in crops present no new safety concerns compared to other breeding practices. The current generation of genome editing technologies is already proving useful to develop new plant varieties with consumer and farmer benefits. Genome editing will likely undergo improved editing specificity along with new developments in SDN delivery and increasing genomic characterization, further improving reagent design and application.
© 2020 American Society of Plant Biologists. All Rights Reserved.
Figures



References
-
- Aflitos S, Schijlen E, de Jong H, de Ridder D, Smit S, Finkers R, Wang J, Zhang G, Li N, Mao L, et al. ; 100 Tomato Genome Sequencing Consortium (2014) Exploring genetic variation in the tomato (Solanum section Lycopersicon) clade by whole-genome sequencing. Plant J 80: 136–148 - PubMed
-
- Aguilar-Meléndez A, Morrell PL, Roose ML, Kim S-C(2009) Genetic diversity and structure in semiwild and domesticated chiles (Capsicum annuum; Solanaceae) from Mexico. Am J Bot 96: 1190–1202 - PubMed
-
- Ahloowalia B, Maluszynski M, Nichterlein K(2004) Global impact of mutation-derived varieties. Euphytica 135: 187–204
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
