

The TRPV3 channel of the bovine rumen: localization and functional characterization of a protein relevant for ruminal ammonia transport
- PMID: 32458085
- PMCID: PMC7293678
- DOI: 10.1007/s00424-020-02393-2
The TRPV3 channel of the bovine rumen: localization and functional characterization of a protein relevant for ruminal ammonia transport
Abstract
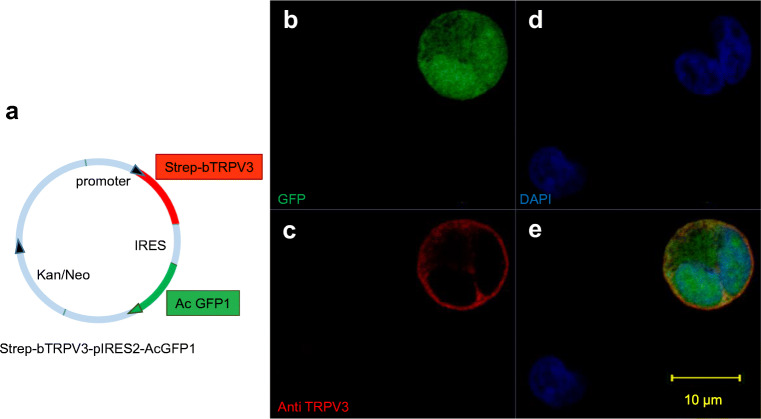
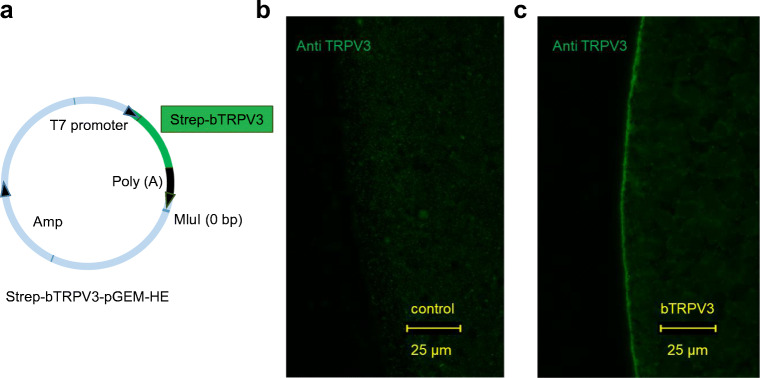
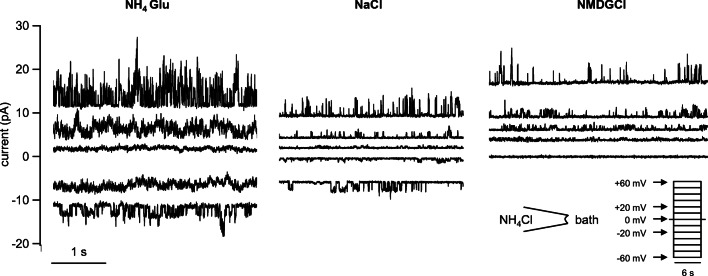
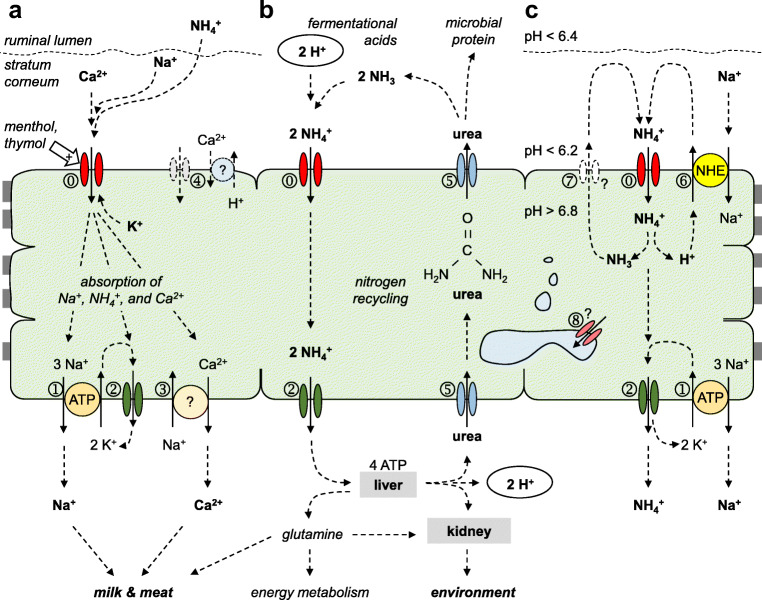
Large quantities of ammonia (NH3 or NH4+) are absorbed from the gut, associated with encephalitis in hepatic disease, poor protein efficiency in livestock, and emissions of nitrogenous climate gasses. Identifying the transport mechanisms appears urgent. Recent functional and mRNA data suggest that absorption of ammonia from the forestomach of cattle may involve TRPV3 channels. The purpose of the present study was to sequence the bovine homologue of TRPV3 (bTRPV3), localize the protein in ruminal tissue, and confirm transport of NH4+. After sequencing, bTRPV3 was overexpressed in HEK-293 cells and Xenopus oocytes. An antibody was selected via epitope screening and used to detect the protein in immunoblots of overexpressing cells and bovine rumen, revealing a signal of the predicted ~ 90 kDa. In rumen only, an additional ~ 60 kDa band appeared, which may represent a previously described bTRPV3 splice variant of equal length. Immunohistochemistry revealed staining from the ruminal stratum basale to stratum granulosum. Measurements with pH-sensitive microelectrodes showed that NH4+ acidifies Xenopus oocytes, with overexpression of bTRPV3 enhancing permeability to NH4+. Single-channel measurements revealed that Xenopus oocytes endogenously expressed small cation channels in addition to fourfold-larger channels only observed after expression of bTRPV3. Both endogenous and bTRPV3 channels conducted NH4+, Na+, and K+. We conclude that bTRPV3 is expressed by the ruminal epithelium on the protein level. In conjunction with data from previous studies, a role in the transport of Na+, Ca2+, and NH4+ emerges. Consequences for calcium homeostasis, ruminal pH, and nitrogen efficiency in cattle are discussed.
Keywords: Ammonia transport; Climate gas; Microelectrode; Rumen; TRPV3; Xenopus oocyte.
Conflict of interest statement
The authors declare that they have no conflicts of interest. Friederike Stumpff is the holder of a patent that was transferred to a startup company.
Figures







Comment in
-
New ways for an old cation.Pflugers Arch. 2020 Jun;472(6):669-670. doi: 10.1007/s00424-020-02394-1. Epub 2020 May 25. Pflugers Arch. 2020. PMID: 32448954 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
