

Base-Pairing Requirements for Small RNA-Mediated Gene Silencing of Recessive Self-Incompatibility Alleles in Arabidopsis halleri
- PMID: 32461267
- PMCID: PMC7337072
- DOI: 10.1534/genetics.120.303351
Base-Pairing Requirements for Small RNA-Mediated Gene Silencing of Recessive Self-Incompatibility Alleles in Arabidopsis halleri
Abstract
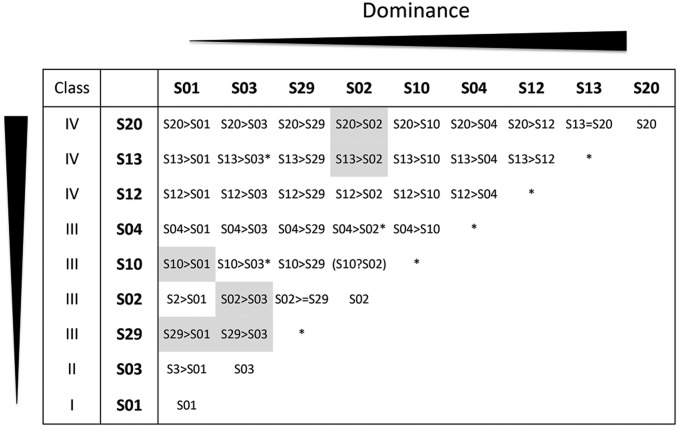
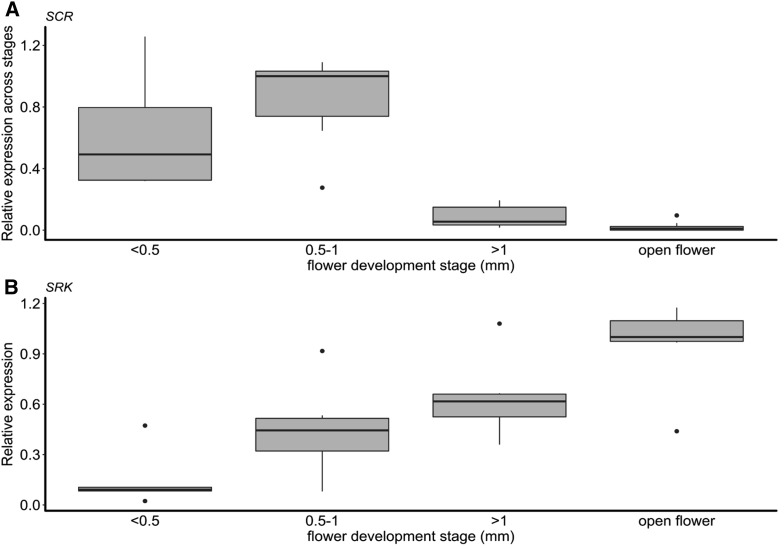
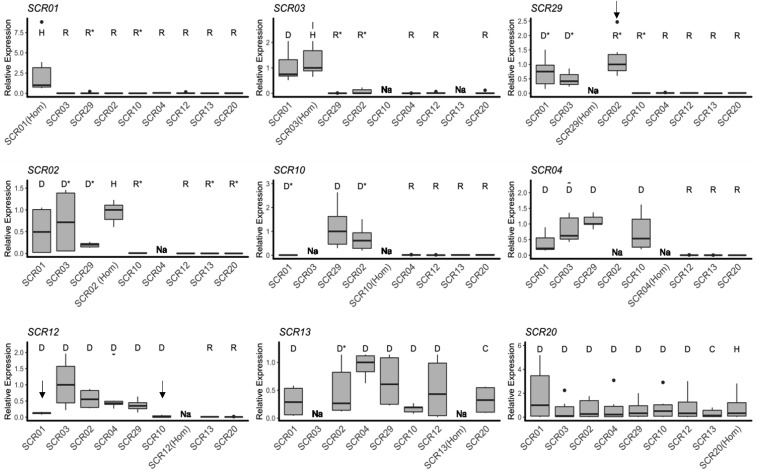
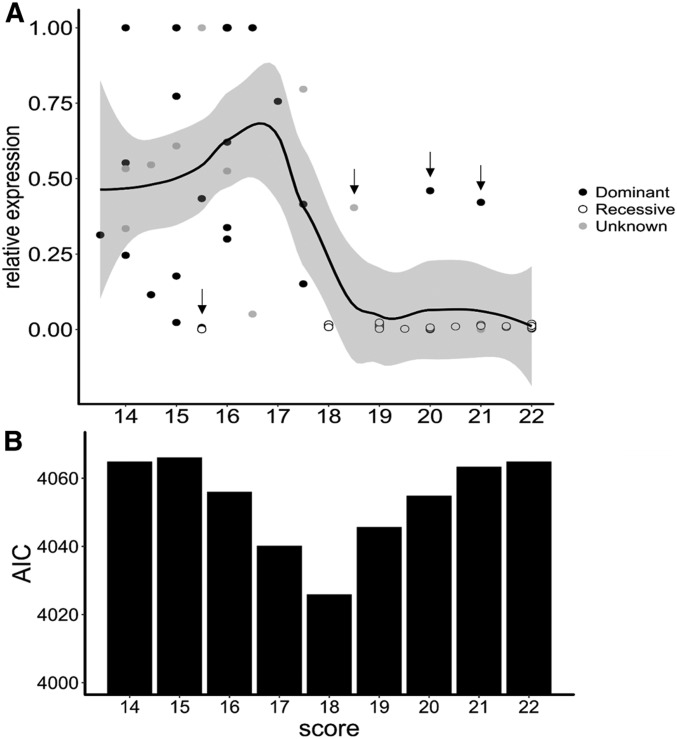
Small noncoding RNAs are central regulators of genome activity and stability. Their regulatory function typically involves sequence similarity with their target sites, but understanding the criteria by which they specifically recognize and regulate their targets across the genome remains a major challenge in the field, especially in the face of the diversity of silencing pathways involved. The dominance hierarchy among self-incompatibility alleles in Brassicaceae is controlled by interactions between a highly diversified set of small noncoding RNAs produced by dominant S-alleles and their corresponding target sites on recessive S-alleles. By controlled crosses, we created numerous heterozygous combinations of S-alleles in Arabidopsis halleri and developed an real-time quantitative PCR assay to compare allele-specific transcript levels for the pollen determinant of self-incompatibility (SCR). This provides the unique opportunity to evaluate the precise base-pairing requirements for effective transcriptional regulation of this target gene. We found strong transcriptional silencing of recessive SCR alleles in all heterozygote combinations examined. A simple threshold model of base pairing for the small RNA-target interaction captures most of the variation in SCR transcript levels. For a subset of S-alleles, we also measured allele-specific transcript levels of the determinant of pistil specificity (SRK), and found sharply distinct expression dynamics throughout flower development between SCR and SRK In contrast to SCR, both SRK alleles were expressed at similar levels in the heterozygote genotypes examined, suggesting no transcriptional control of dominance for this gene. We discuss the implications for the evolutionary processes associated with the origin and maintenance of the dominance hierarchy among self-incompatibility alleles.
Keywords: Arabidopsis halleri; RT-qPCR; allele-specific expression assay; dominance/recessivity; sporophytic self-incompatibility.
Copyright © 2020 by the Genetics Society of America.
Figures

References
-
- Bates D., Mächler M., Bolker B., and Walker S., 2014. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67: 1–48.
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Research Materials
