

FcRn, but not FcγRs, drives maternal-fetal transplacental transport of human IgG antibodies
- PMID: 32461366
- PMCID: PMC7293622
- DOI: 10.1073/pnas.2004325117
FcRn, but not FcγRs, drives maternal-fetal transplacental transport of human IgG antibodies
Abstract
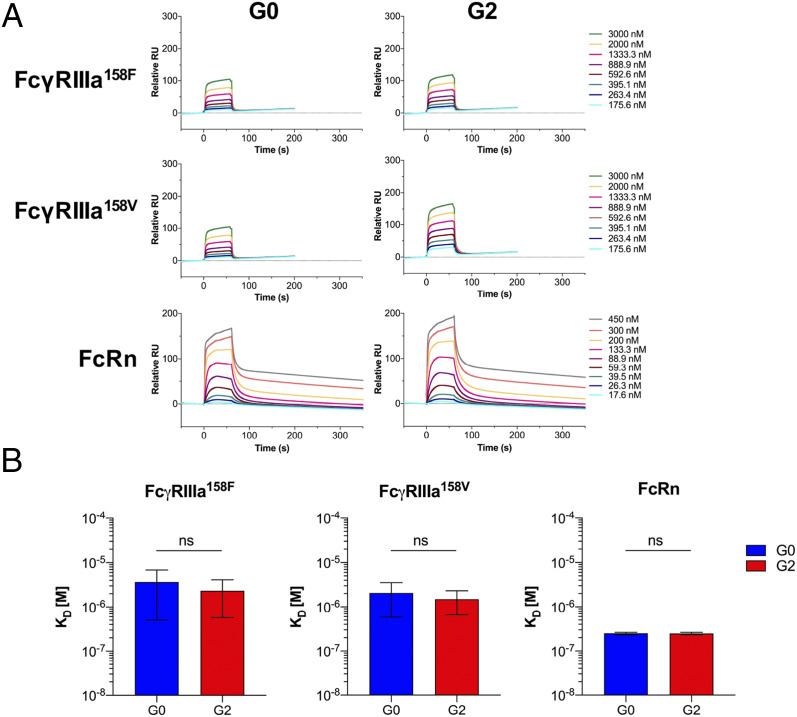
The IgG Fc domain has the capacity to interact with diverse types of receptors, including the neonatal Fc receptor (FcRn) and Fcγ receptors (FcγRs), which confer pleiotropic biological activities. Whereas FcRn regulates IgG epithelial transport and recycling, Fc effector activities, such as antibody-dependent cellular cytotoxicity (ADCC) and phagocytosis, are mediated by FcγRs, which upon cross-linking transduce signals that modulate the function of effector leukocytes. Despite the well-defined and nonoverlapping functional properties of FcRn and FcγRs, recent studies have suggested that FcγRs mediate transplacental IgG transport, as certain Fc glycoforms were reported to be enriched in fetal circulation. To determine the contribution of FcγRs and FcRn to the maternal-fetal transport of IgG, we characterized the IgG Fc glycosylation in paired maternal-fetal samples from patient cohorts from Uganda and Nicaragua. No differences in IgG1 Fc glycan profiles and minimal differences in IgG2 Fc glycans were noted, whereas the presence or absence of galactose on the Fc glycan of IgG1 did not alter FcγRIIIa or FcRn binding, half-life, or their ability to deplete target cells in FcγR/FcRn humanized mice. Modeling maternal-fetal transport in FcγR/FcRn humanized mice confirmed that only FcRn contributed to transplacental transport of IgG; IgG selectively enhanced for FcRn binding resulted in enhanced accumulation of maternal antibody in the fetus. In contrast, enhancing FcγRIIIa binding did not result in enhanced maternal-fetal transport. These results argue against a role for FcγRs in IgG transplacental transport, suggesting Fc engineering of maternally administered antibody to enhance only FcRn binding as a means to improve maternal-fetal transport of IgG.
Keywords: Fc domain; FcRn; IgG; immunoglobulin; placental transfer.
Conflict of interest statement
The authors declare no competing interest.
Figures






Similar articles
-
Reduced FcRn-mediated transcytosis of IgG2 due to a missing Glycine in its lower hinge.Sci Rep. 2019 May 14;9(1):7363. doi: 10.1038/s41598-019-40731-2. Sci Rep. 2019. PMID: 31089170 Free PMC article.
-
Combined glyco- and protein-Fc engineering simultaneously enhance cytotoxicity and half-life of a therapeutic antibody.MAbs. 2014 Mar-Apr;6(2):422-36. doi: 10.4161/mabs.27854. Epub 2014 Jan 15. MAbs. 2014. PMID: 24492301 Free PMC article.
-
Human IgG lacking effector functions demonstrate lower FcRn-binding and reduced transplacental transport.Mol Immunol. 2018 Mar;95:1-9. doi: 10.1016/j.molimm.2018.01.006. Epub 2018 Feb 20. Mol Immunol. 2018. PMID: 29367080
-
Relevance of the Materno-Fetal Interface for the Induction of Antigen-Specific Immune Tolerance.Front Immunol. 2020 May 14;11:810. doi: 10.3389/fimmu.2020.00810. eCollection 2020. Front Immunol. 2020. PMID: 32477339 Free PMC article. Review.
-
The Neonatal Fc Receptor (FcRn): A Misnomer?Front Immunol. 2019 Jul 10;10:1540. doi: 10.3389/fimmu.2019.01540. eCollection 2019. Front Immunol. 2019. PMID: 31354709 Free PMC article. Review.
Cited by
-
FcRn-enhancing mutations lead to increased and prolonged levels of the HIV CCR5-blocking monoclonal antibody leronlimab in the fetuses and newborns of pregnant rhesus macaques.MAbs. 2024 Jan-Dec;16(1):2406788. doi: 10.1080/19420862.2024.2406788. Epub 2024 Sep 26. MAbs. 2024. PMID: 39324549 Free PMC article.
-
Mechanism of glycoform specificity and in vivo protection by an anti-afucosylated IgG nanobody.Nat Commun. 2023 May 18;14(1):2853. doi: 10.1038/s41467-023-38453-1. Nat Commun. 2023. PMID: 37202422 Free PMC article.
-
Evaluation of transplacental transfer of mRNA vaccine products and functional antibodies during pregnancy and early infancy.Res Sq [Preprint]. 2021 Dec 15:rs.3.rs-1150427. doi: 10.21203/rs.3.rs-1150427/v1. Res Sq. 2021. Update in: Nat Commun. 2022 Jul 30;13(1):4422. doi: 10.1038/s41467-022-32188-1. PMID: 34931183 Free PMC article. Updated. Preprint.
-
HIV-related Differences in Placental Immunology: Data From the PRACHITi Cohort in Pune, India.Open Forum Infect Dis. 2025 Jan 31;12(3):ofaf047. doi: 10.1093/ofid/ofaf047. eCollection 2025 Mar. Open Forum Infect Dis. 2025. PMID: 40046890 Free PMC article.
-
Antibody Immunity to Zika Virus among Young Children in a Flavivirus-Endemic Area in Nicaragua.Viruses. 2023 Mar 21;15(3):796. doi: 10.3390/v15030796. Viruses. 2023. PMID: 36992504 Free PMC article.
References
-
- Firan M. et al. ., The MHC class I-related receptor, FcRn, plays an essential role in the maternofetal transfer of gamma-globulin in humans. Int. Immunol. 13, 993–1002 (2001). - PubMed
-
- Ghetie V., Ward E. S., FcRn: The MHC class I-related receptor that is more than an IgG transporter. Immunol. Today 18, 592–598 (1997). - PubMed
-
- Simister N. E., Jacobowitz Israel E., Ahouse J. C., Story C. M., New functions of the MHC class I-related Fc receptor, FcRn. Biochem. Soc. Trans. 25, 481–486 (1997). - PubMed
-
- Shields R. L. et al. ., High resolution mapping of the binding site on human IgG1 for Fc gamma RI, Fc gamma RII, Fc gamma RIII, and FcRn and design of IgG1 variants with improved binding to the Fc gamma R. J. Biol. Chem. 276, 6591–6604 (2001). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
