

Prc1-rich kinetochores are required for error-free acentrosomal spindle bipolarization during meiosis I in mouse oocytes
- PMID: 32461611
- PMCID: PMC7253481
- DOI: 10.1038/s41467-020-16488-y
Prc1-rich kinetochores are required for error-free acentrosomal spindle bipolarization during meiosis I in mouse oocytes
Abstract
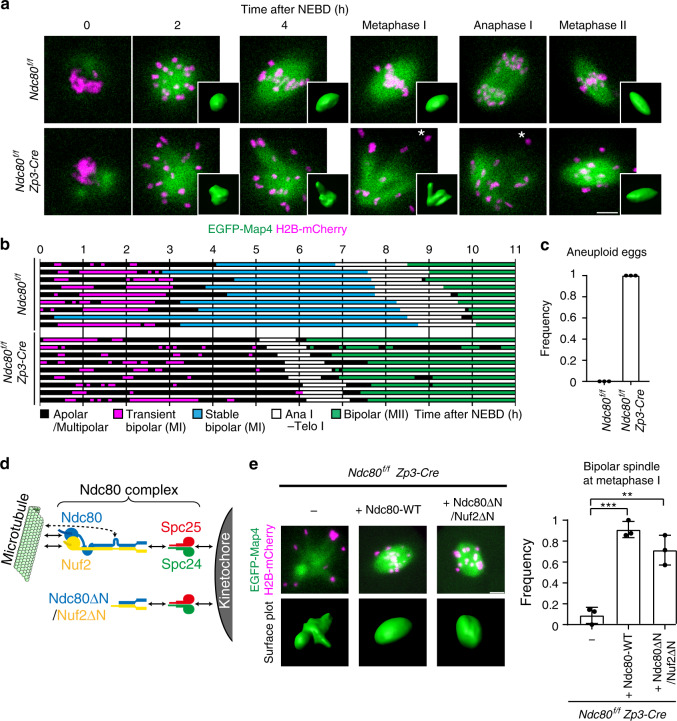
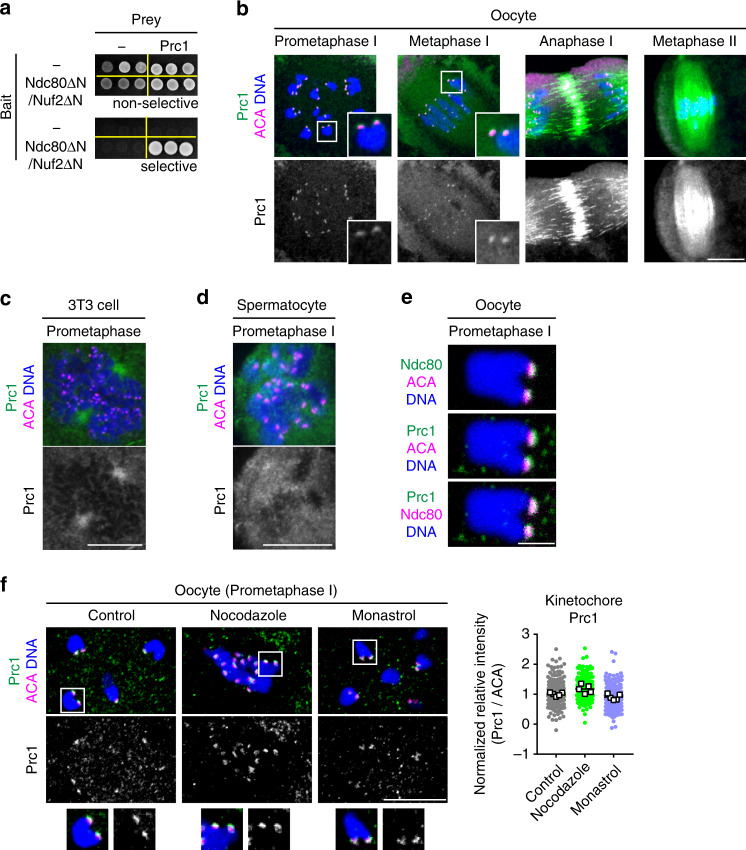
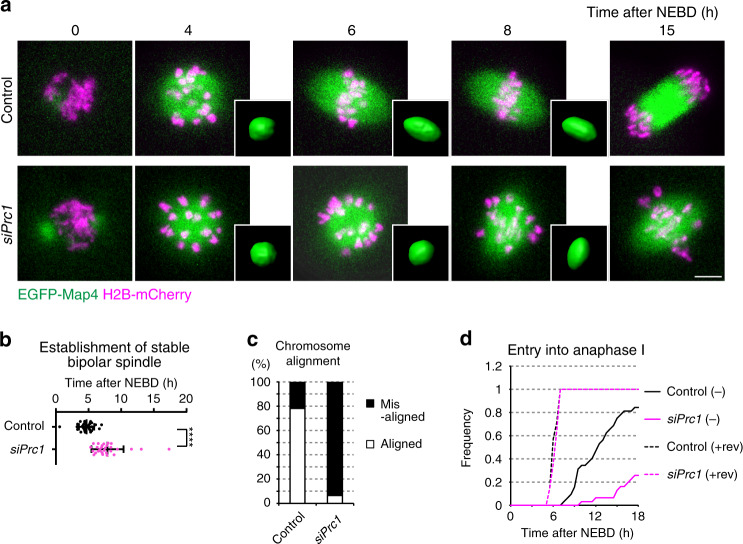
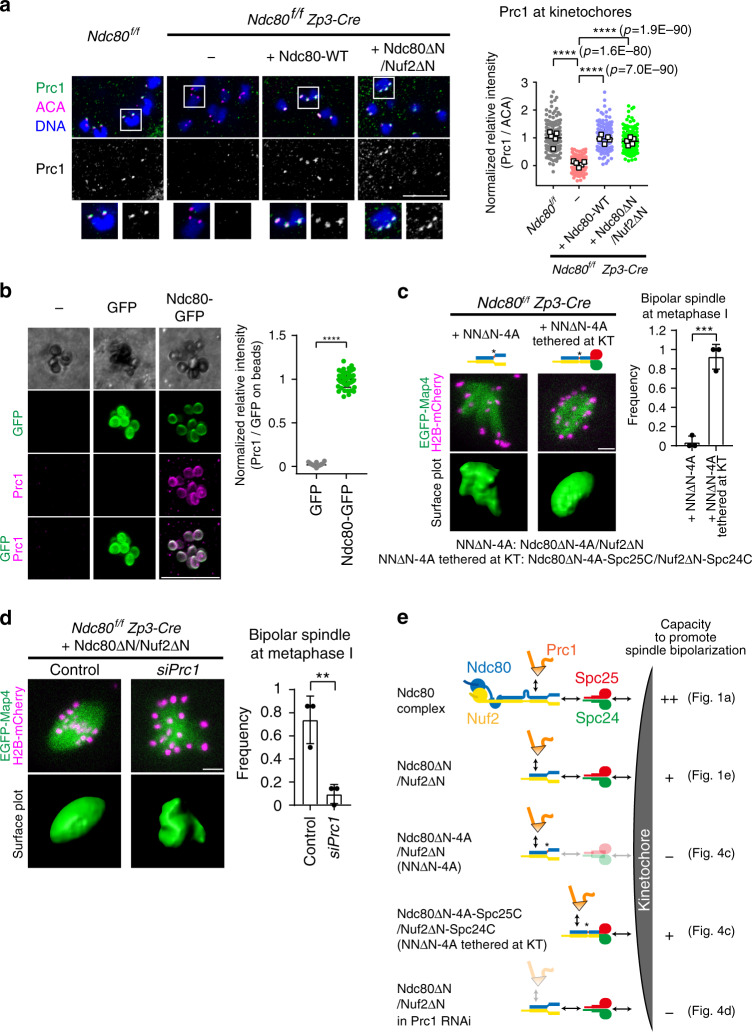
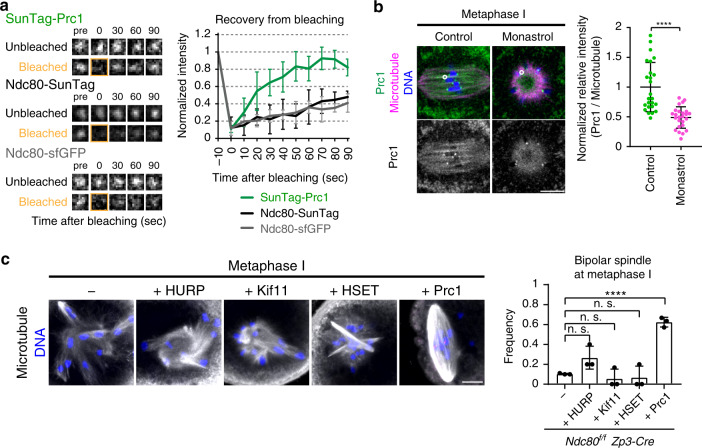
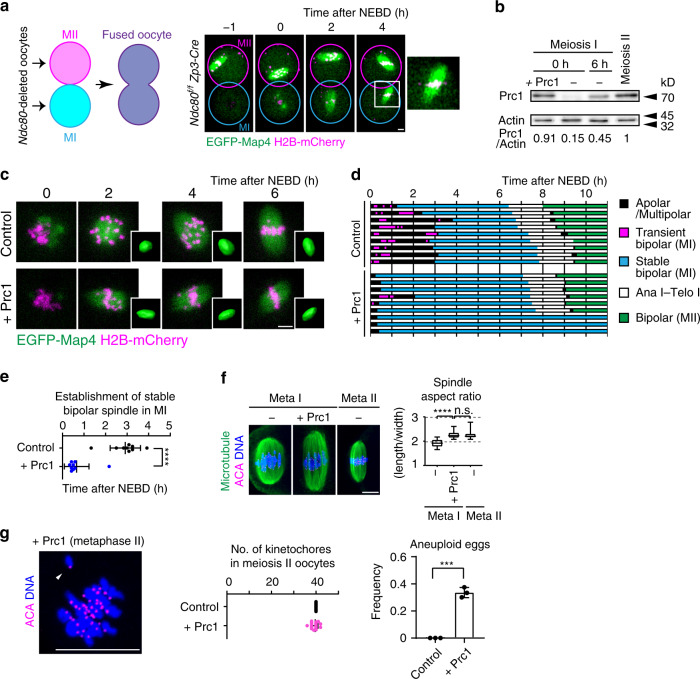
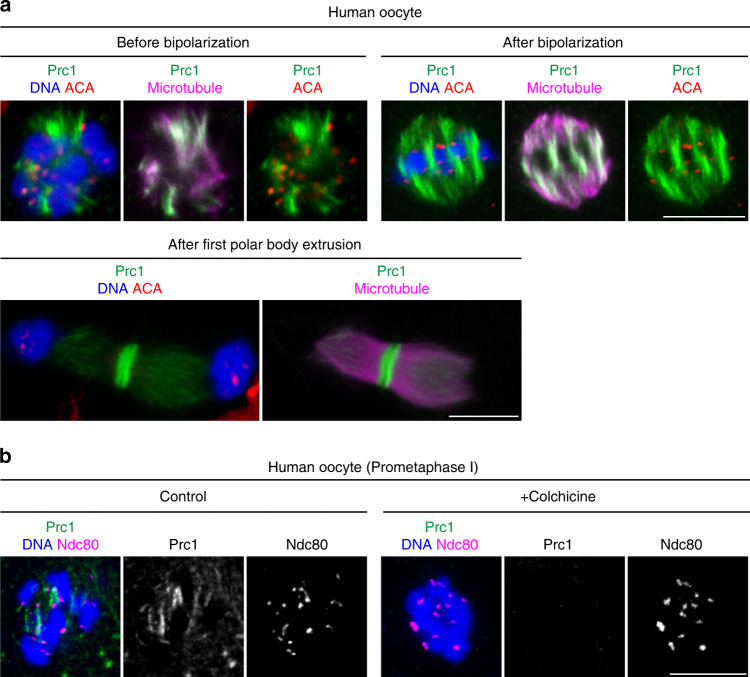
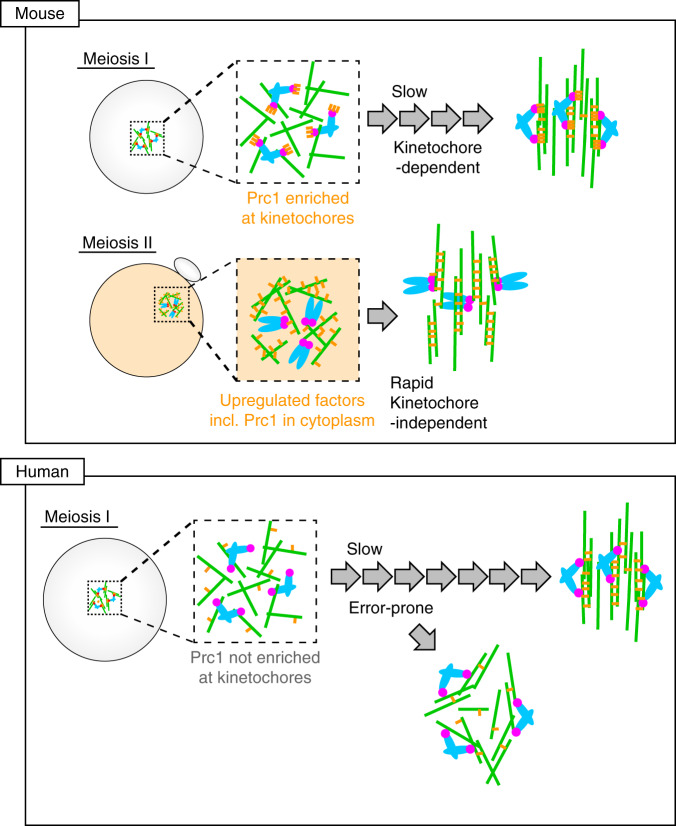
Acentrosomal meiosis in oocytes represents a gametogenic challenge, requiring spindle bipolarization without predefined bipolar cues. While much is known about the structures that promote acentrosomal microtubule nucleation, less is known about the structures that mediate spindle bipolarization in mammalian oocytes. Here, we show that in mouse oocytes, kinetochores are required for spindle bipolarization in meiosis I. This process is promoted by oocyte-specific, microtubule-independent enrichment of the antiparallel microtubule crosslinker Prc1 at kinetochores via the Ndc80 complex. In contrast, in meiosis II, cytoplasm that contains upregulated factors including Prc1 supports kinetochore-independent pathways for spindle bipolarization. The kinetochore-dependent mode of spindle bipolarization is required for meiosis I to prevent chromosome segregation errors. Human oocytes, where spindle bipolarization is reportedly error prone, exhibit no detectable kinetochore enrichment of Prc1. This study reveals an oocyte-specific function of kinetochores in acentrosomal spindle bipolarization in mice, and provides insights into the error-prone nature of human oocytes.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Cdk1 negatively regulates the spindle localization of Prc1 in mouse oocytes.Genes Cells. 2020 Oct;25(10):685-694. doi: 10.1111/gtc.12803. Epub 2020 Sep 21. Genes Cells. 2020. PMID: 32865279
-
MPS1 promotes timely spindle bipolarization to prevent kinetochore-microtubule attachment errors in oocytes.EMBO J. 2025 Jul;44(13):3794-3823. doi: 10.1038/s44318-025-00461-w. Epub 2025 Jun 4. EMBO J. 2025. PMID: 40467861 Free PMC article.
-
Mechanisms of minor pole-mediated spindle bipolarization in human oocytes.Science. 2024 Aug 23;385(6711):eado1022. doi: 10.1126/science.ado1022. Epub 2024 Aug 23. Science. 2024. PMID: 39172836
-
Meiotic defects in human oocytes: Potential causes and clinical implications.Bioessays. 2022 Dec;44(12):e2200135. doi: 10.1002/bies.202200135. Epub 2022 Oct 7. Bioessays. 2022. PMID: 36207289 Review.
-
Aneuploidy in human eggs: contributions of the meiotic spindle.Biochem Soc Trans. 2021 Feb 26;49(1):107-118. doi: 10.1042/BST20200043. Biochem Soc Trans. 2021. PMID: 33449109 Free PMC article. Review.
Cited by
-
Evidence for anaphase pulling forces during C. elegans meiosis.J Cell Biol. 2020 Dec 7;219(12):e202005179. doi: 10.1083/jcb.202005179. J Cell Biol. 2020. PMID: 33064834 Free PMC article.
-
N-Terminus-Mediated Solution Structure of Dimerization Domain of PRC1.Curr Issues Mol Biol. 2022 Apr 10;44(4):1626-1645. doi: 10.3390/cimb44040111. Curr Issues Mol Biol. 2022. PMID: 35723369 Free PMC article.
-
Proteome of oocyte spindle identifies Ccdc69 regulates spindle assembly like "band-tightening spell".Cell Mol Life Sci. 2025 Jul 30;82(1):292. doi: 10.1007/s00018-025-05821-7. Cell Mol Life Sci. 2025. PMID: 40736675 Free PMC article.
-
Kinetochore- and chromosome-driven transition of microtubules into bundles promotes spindle assembly.Nat Commun. 2022 Nov 27;13(1):7307. doi: 10.1038/s41467-022-34957-4. Nat Commun. 2022. PMID: 36435852 Free PMC article.
-
Validation of the solution structure of dimerization domain of PRC1.PLoS One. 2022 Aug 5;17(8):e0270572. doi: 10.1371/journal.pone.0270572. eCollection 2022. PLoS One. 2022. PMID: 35930764 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
