

Hepatic NADH reductive stress underlies common variation in metabolic traits
- PMID: 32461692
- PMCID: PMC7536642
- DOI: 10.1038/s41586-020-2337-2
Hepatic NADH reductive stress underlies common variation in metabolic traits
Abstract
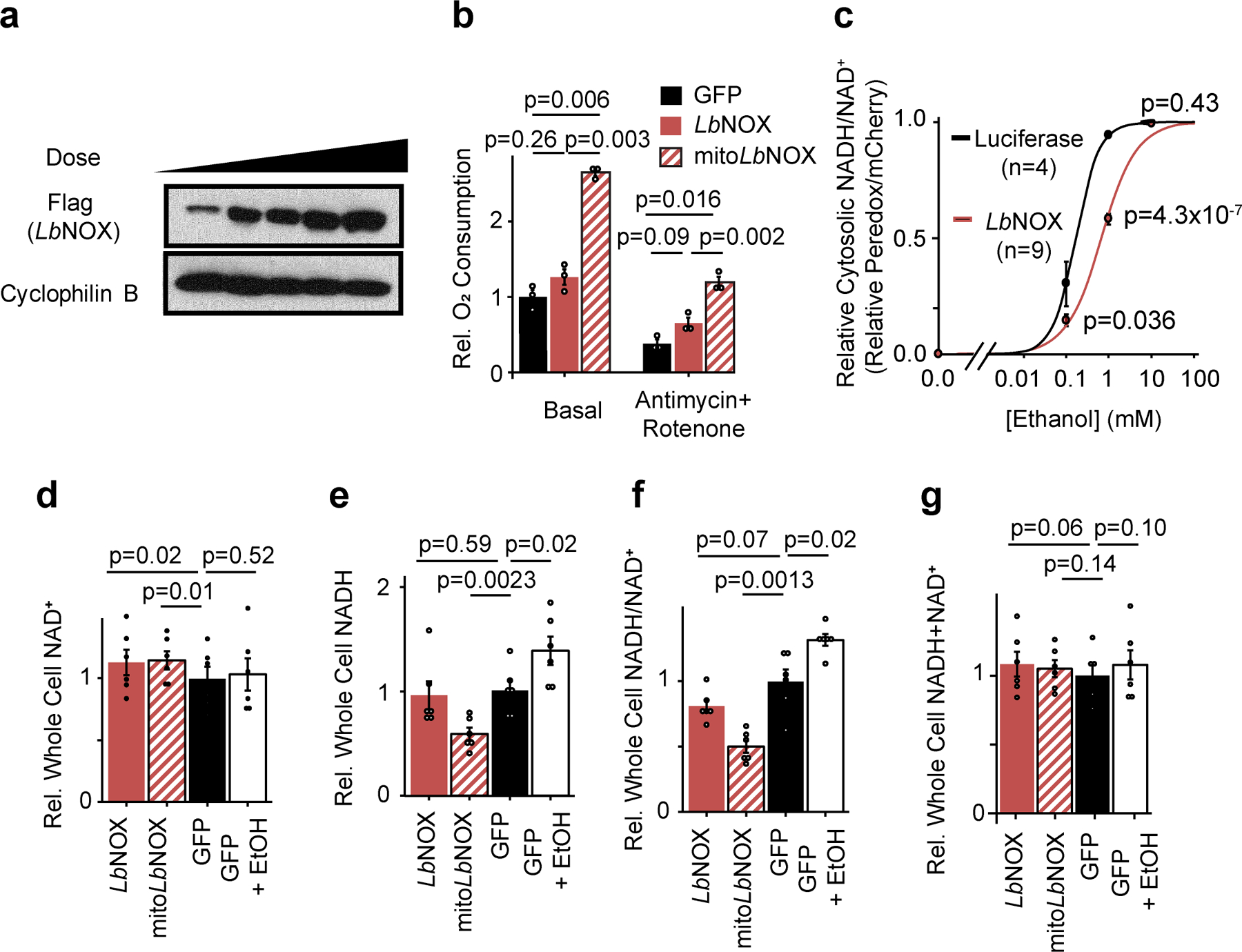
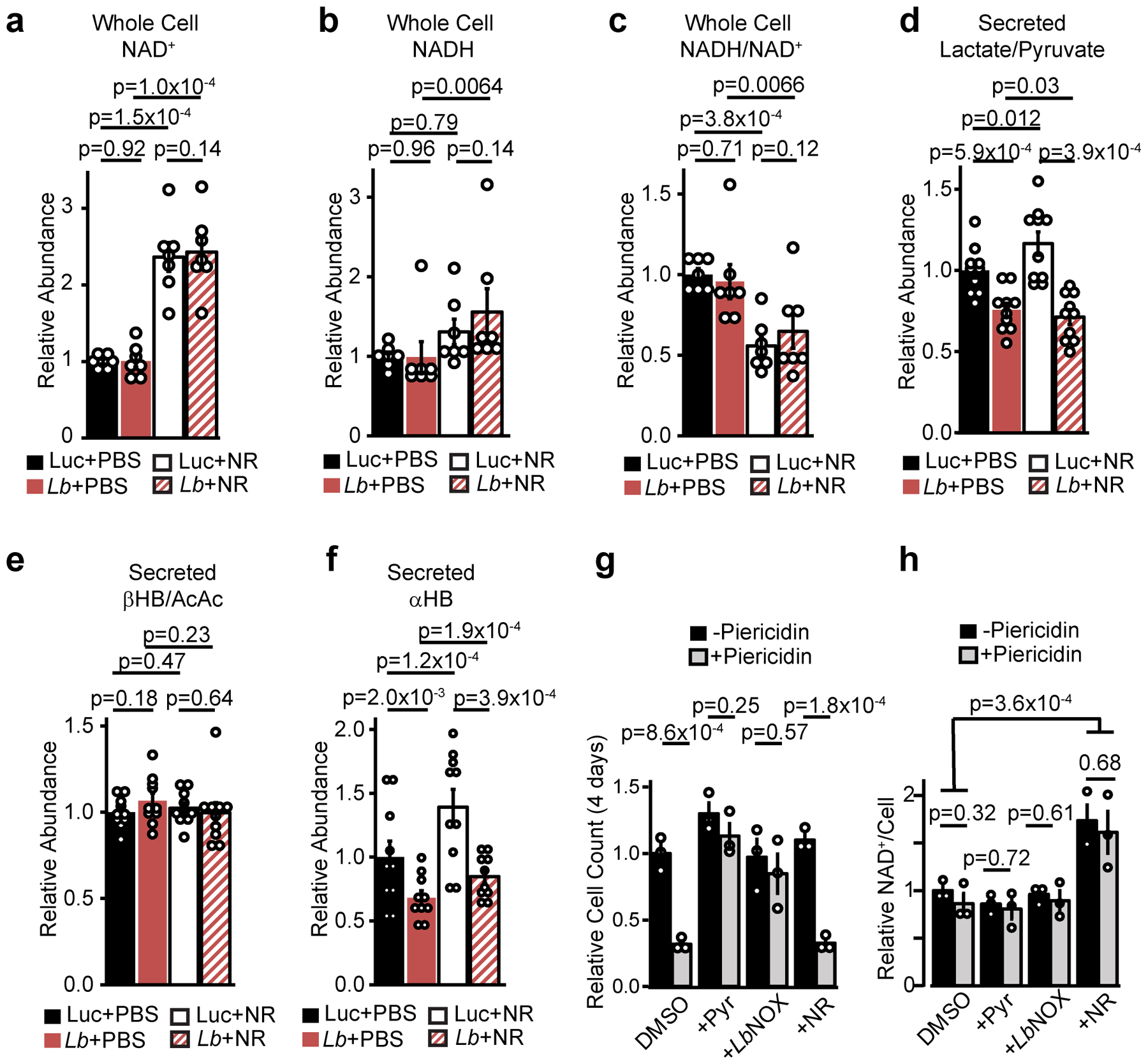
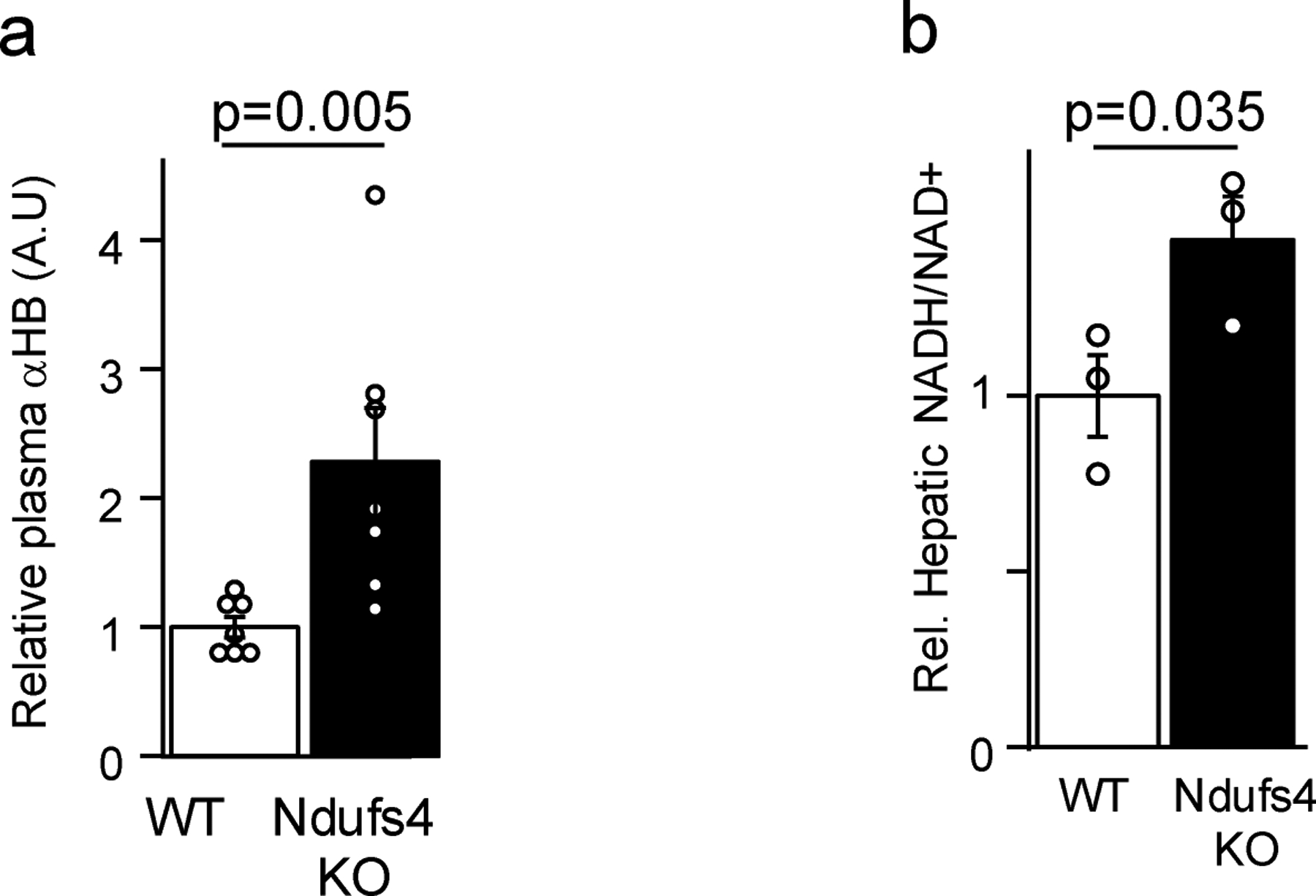
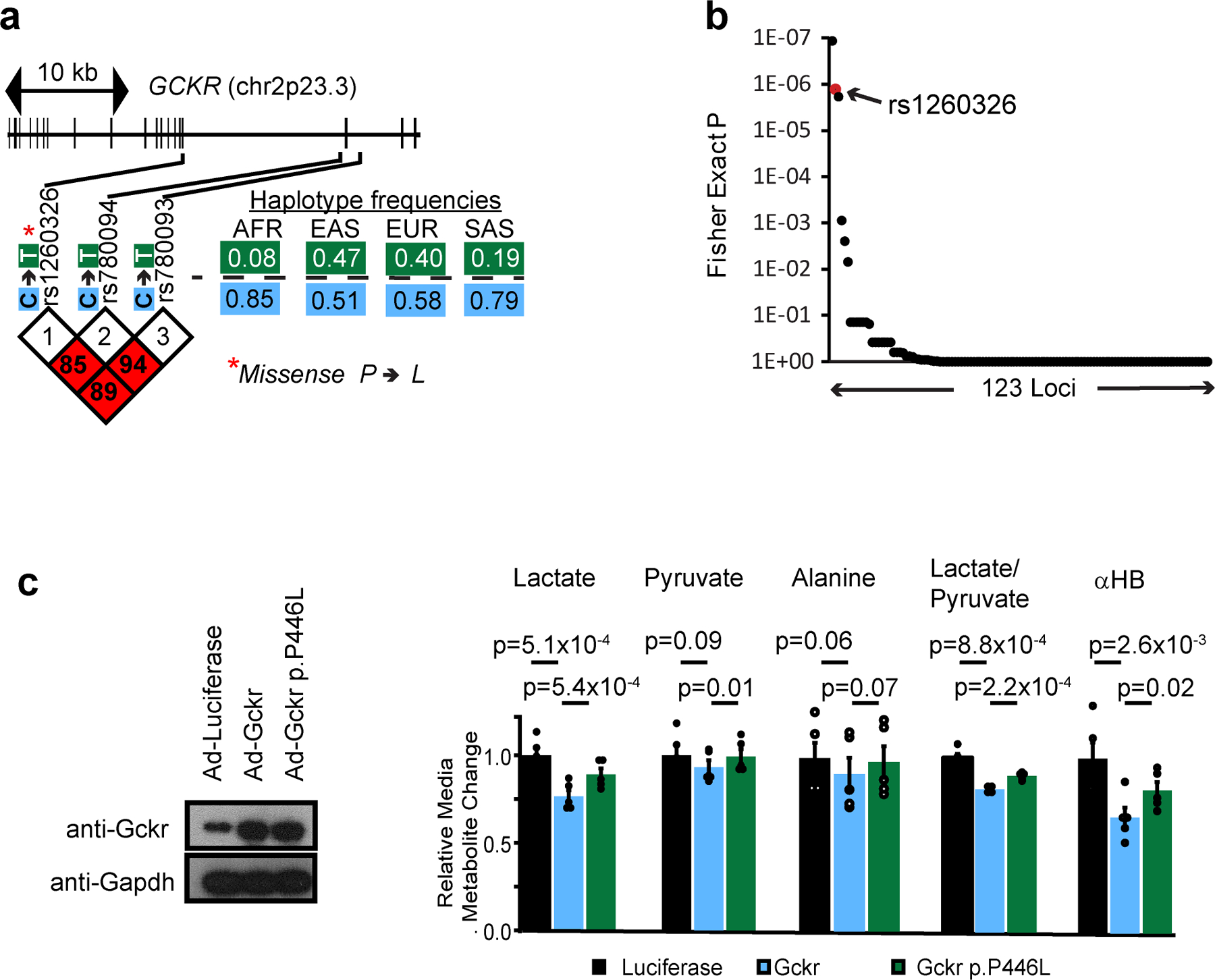
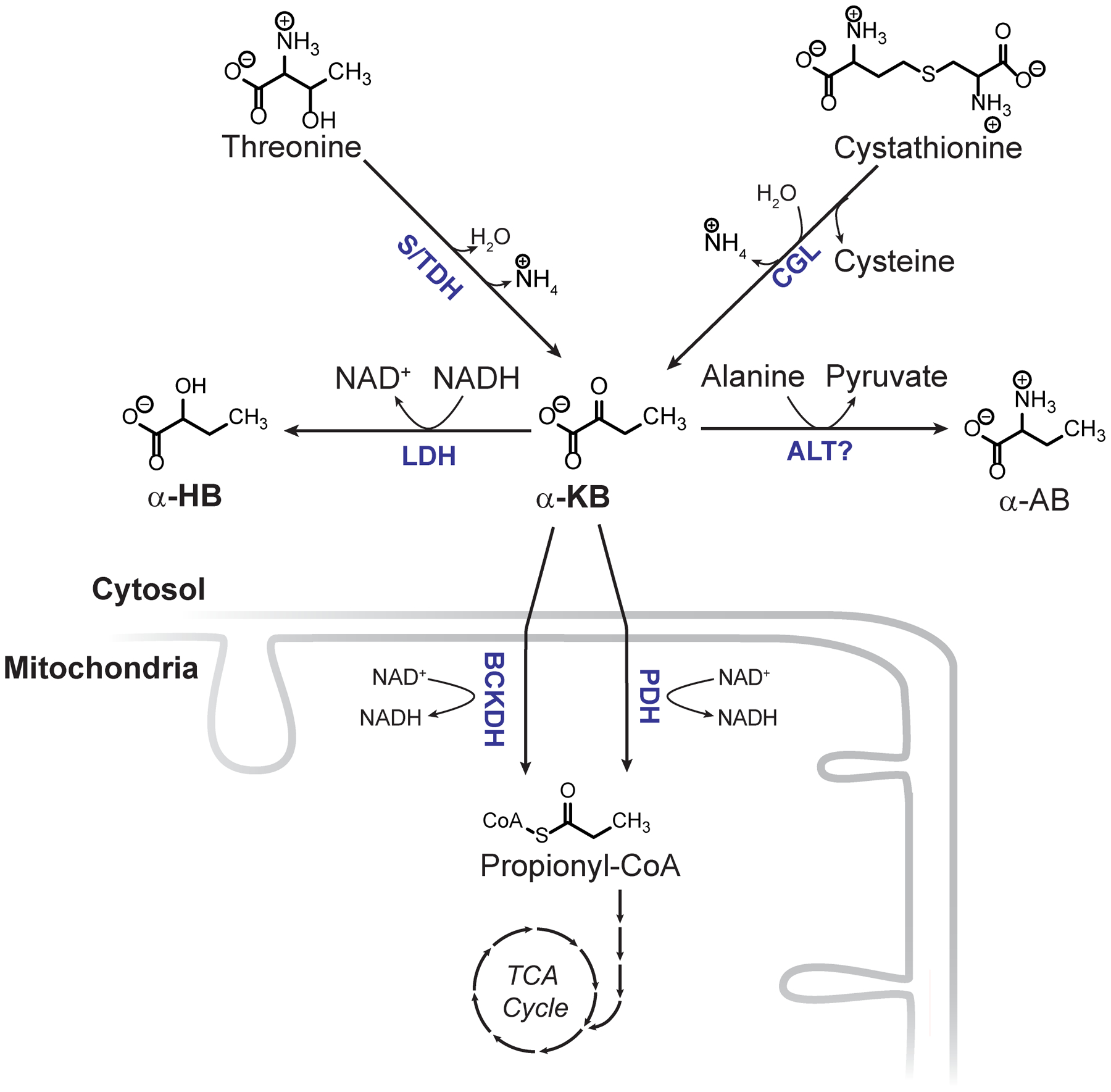
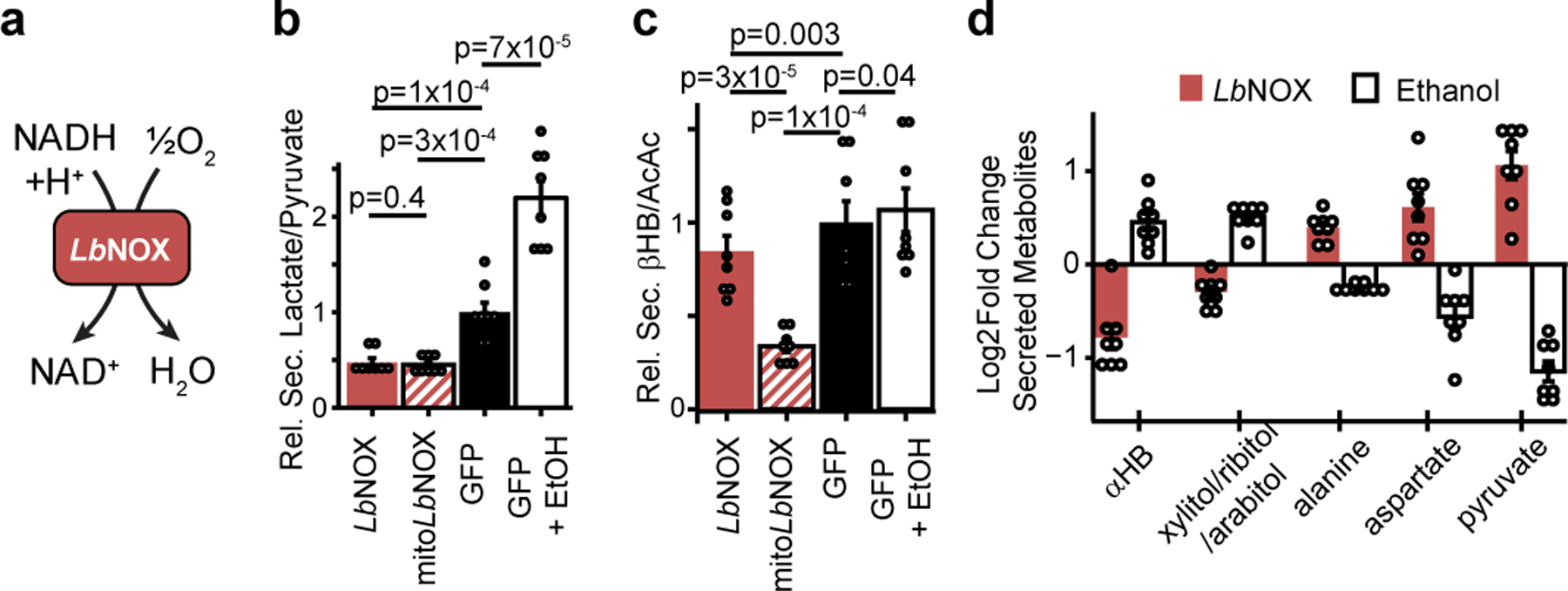
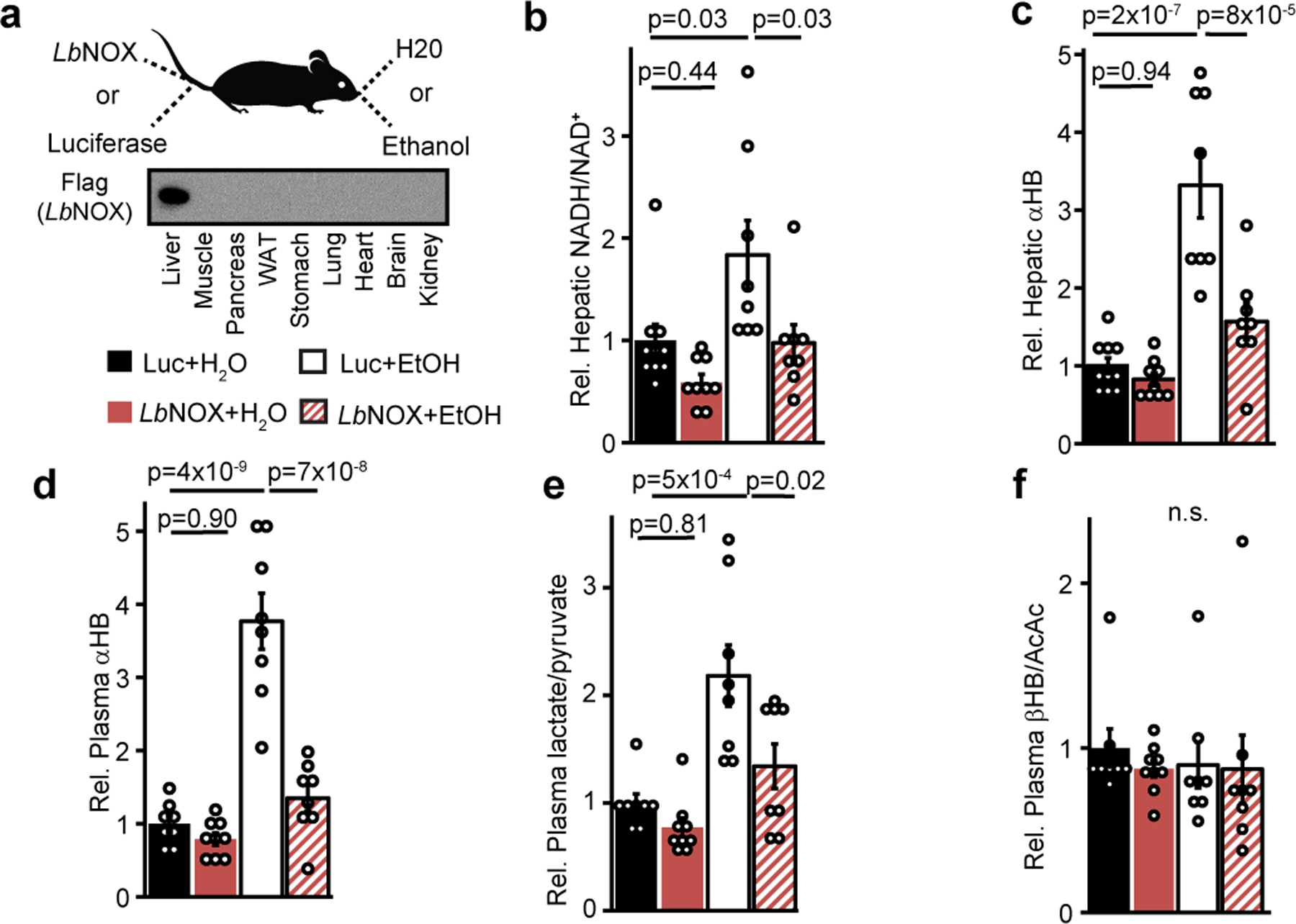
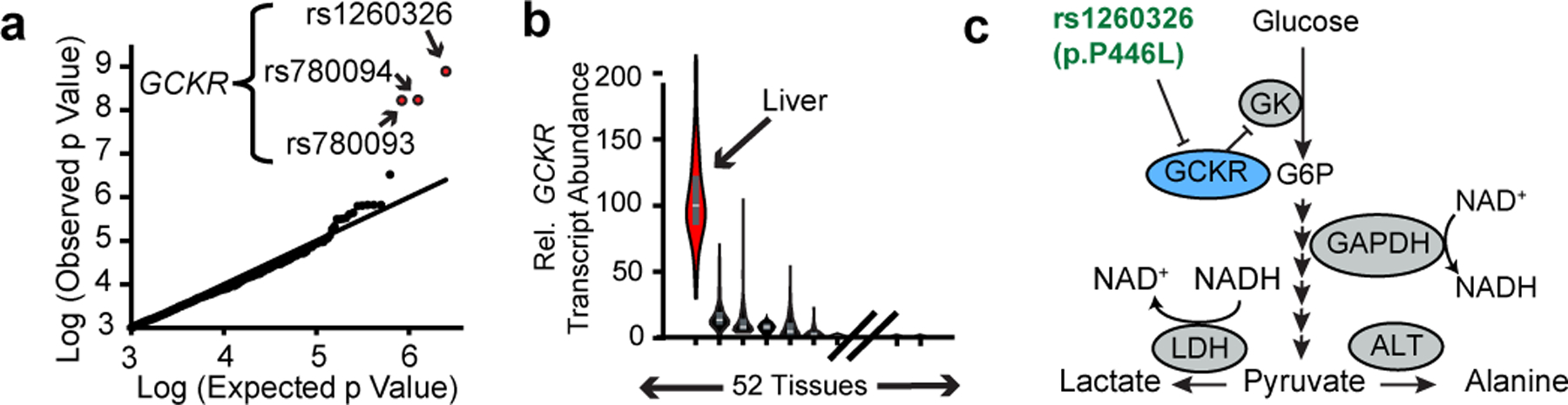
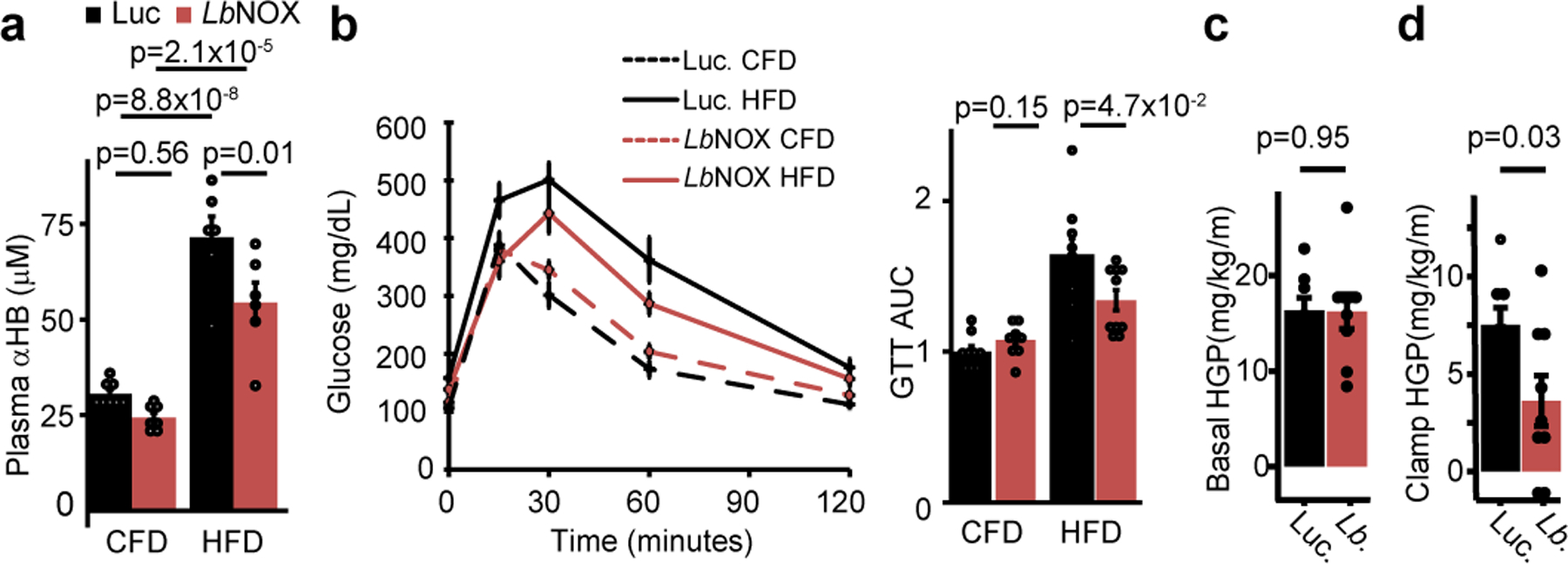
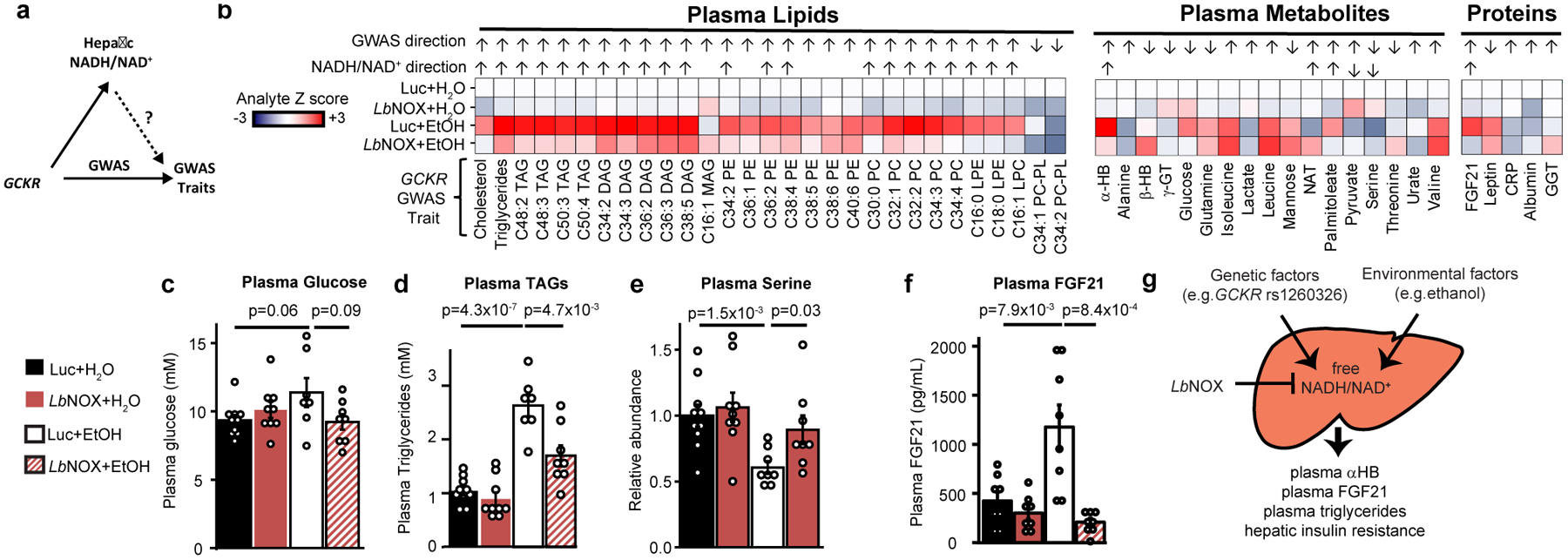
The cellular NADH/NAD+ ratio is fundamental to biochemistry, but the extent to which it reflects versus drives metabolic physiology in vivo is poorly understood. Here we report the in vivo application of Lactobacillus brevis (Lb)NOX1, a bacterial water-forming NADH oxidase, to assess the metabolic consequences of directly lowering the hepatic cytosolic NADH/NAD+ ratio in mice. By combining this genetic tool with metabolomics, we identify circulating α-hydroxybutyrate levels as a robust marker of an elevated hepatic cytosolic NADH/NAD+ ratio, also known as reductive stress. In humans, elevations in circulating α-hydroxybutyrate levels have previously been associated with impaired glucose tolerance2, insulin resistance3 and mitochondrial disease4, and are associated with a common genetic variant in GCKR5, which has previously been associated with many seemingly disparate metabolic traits. Using LbNOX, we demonstrate that NADH reductive stress mediates the effects of GCKR variation on many metabolic traits, including circulating triglyceride levels, glucose tolerance and FGF21 levels. Our work identifies an elevated hepatic NADH/NAD+ ratio as a latent metabolic parameter that is shaped by human genetic variation and contributes causally to key metabolic traits and diseases. Moreover, it underscores the utility of genetic tools such as LbNOX to empower studies of 'causal metabolism'.
Conflict of interest statement
Figures
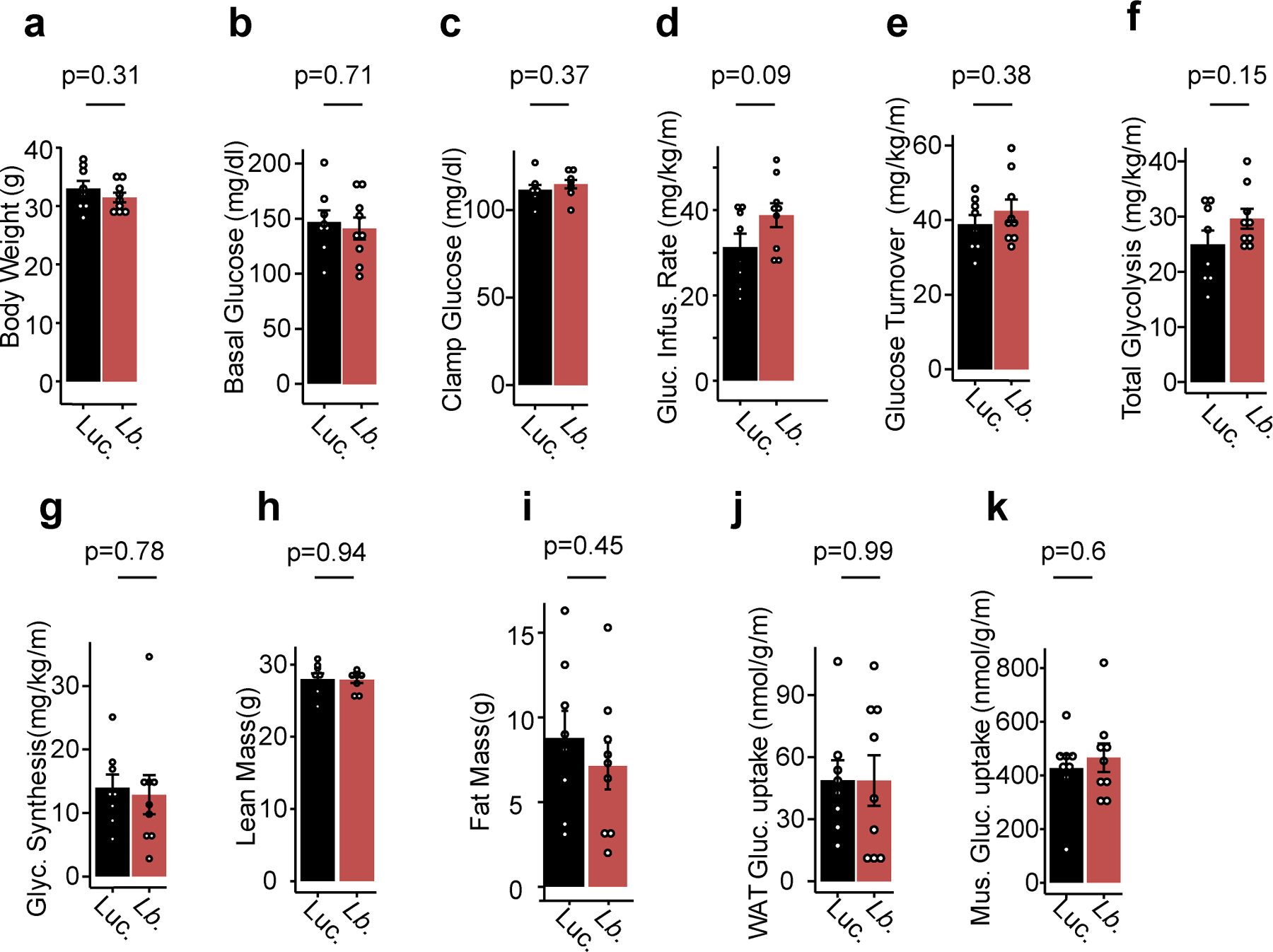
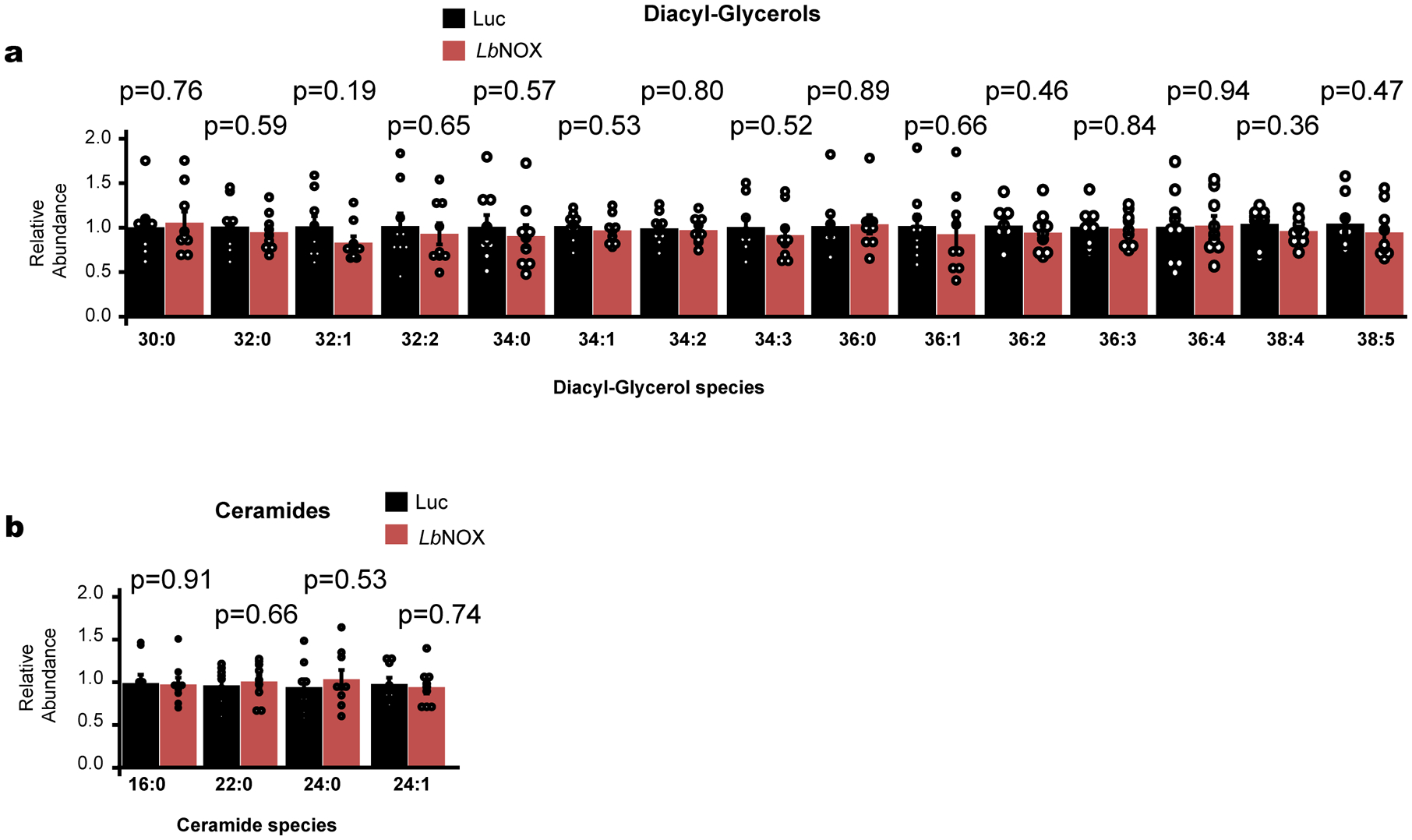
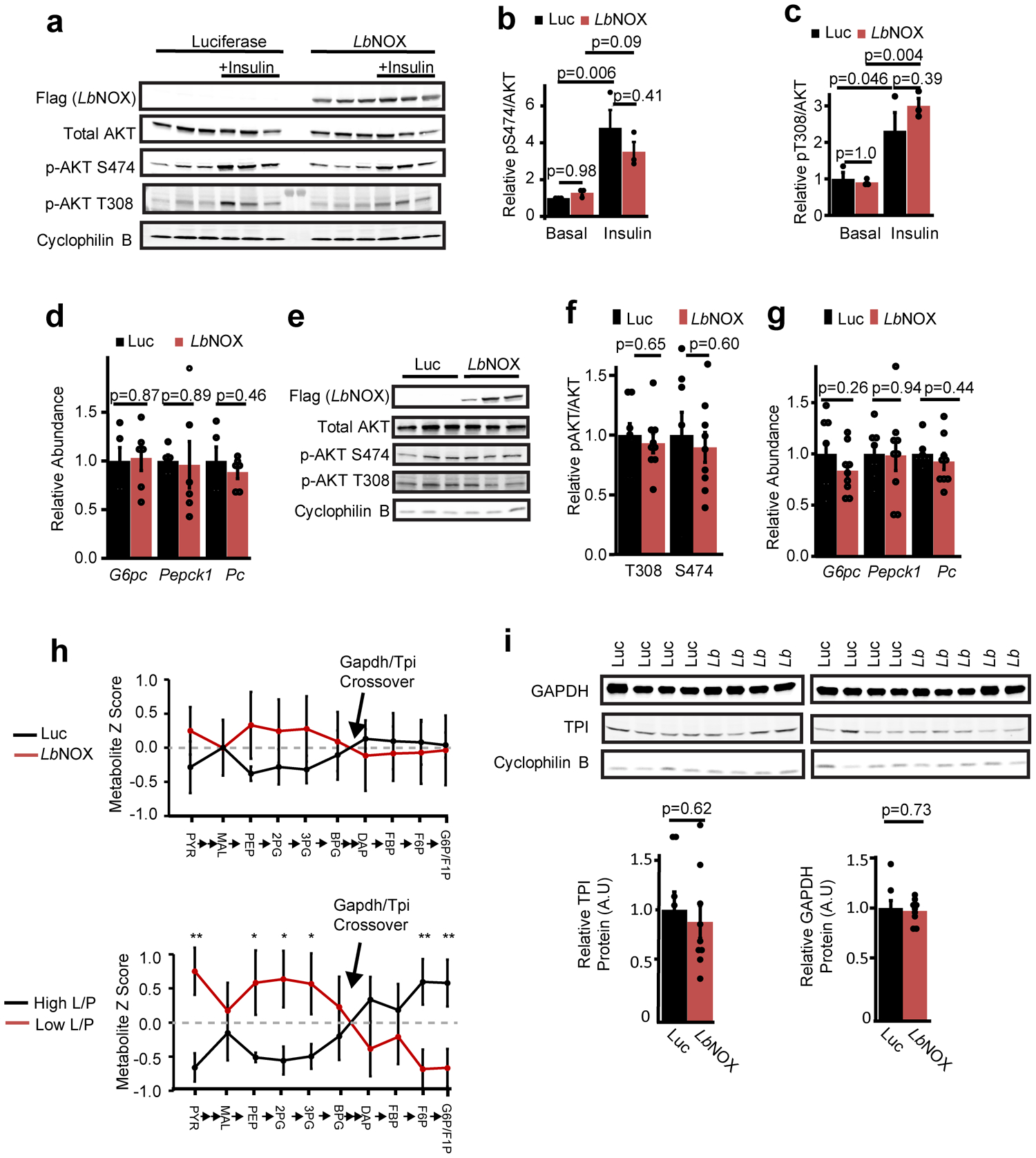
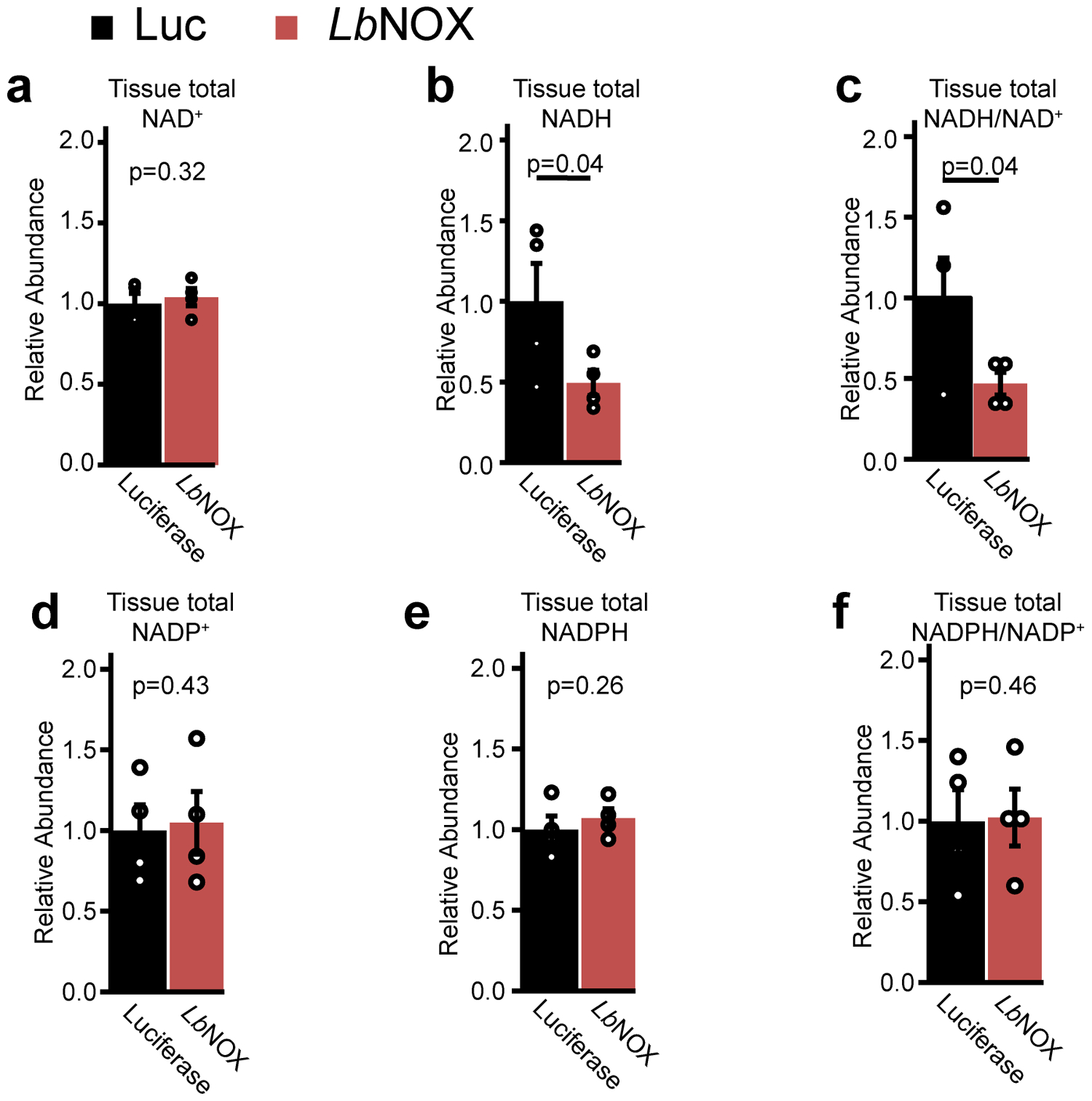
References
-
- Cobb J et al. α-Hydroxybutyric acid is a selective metabolite biomarker of impaired glucose tolerance. Diabetes Care 39, 988–995 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 GM099683/GM/NIGMS NIH HHS/United States
- P30 DK040561/DK/NIDDK NIH HHS/United States
- R01 GM124038/GM/NIGMS NIH HHS/United States
- 5U2C-DK093000/NH/NIH HHS/United States
- P30 DK043351/DK/NIDDK NIH HHS/United States
- R35GM122455/NH/NIH HHS/United States
- K08DK1158811/NH/NIH HHS/United States
- R35 GM122455/GM/NIGMS NIH HHS/United States
- T32 DK110919/DK/NIDDK NIH HHS/United States
- TR01GM099683/NH/NIH HHS/United States
- DP1 EB016985/EB/NIBIB NIH HHS/United States
- K08 DK115881/DK/NIDDK NIH HHS/United States
- F32 GM133047/GM/NIGMS NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- F32 DK111132/DK/NIDDK NIH HHS/United States
- T32 HG002295/HG/NHGRI NIH HHS/United States
- R01 DK075787/DK/NIDDK NIH HHS/United States
- R01 DK134675/DK/NIDDK NIH HHS/United States
- U2C DK093000/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
