

The protective role of m1A during stress-induced granulation
- PMID: 32462207
- PMCID: PMC7883823
- DOI: 10.1093/jmcb/mjaa023
The protective role of m1A during stress-induced granulation
Abstract
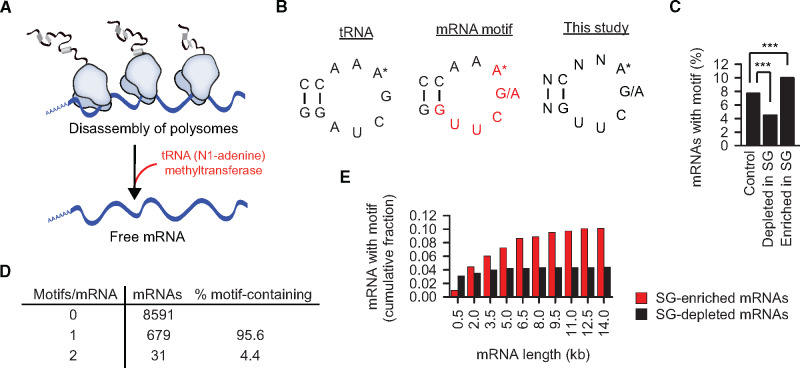
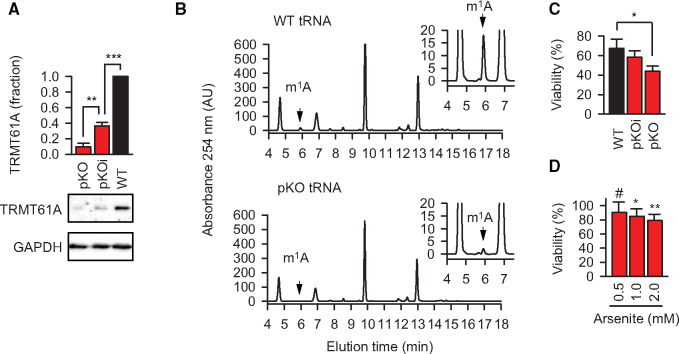
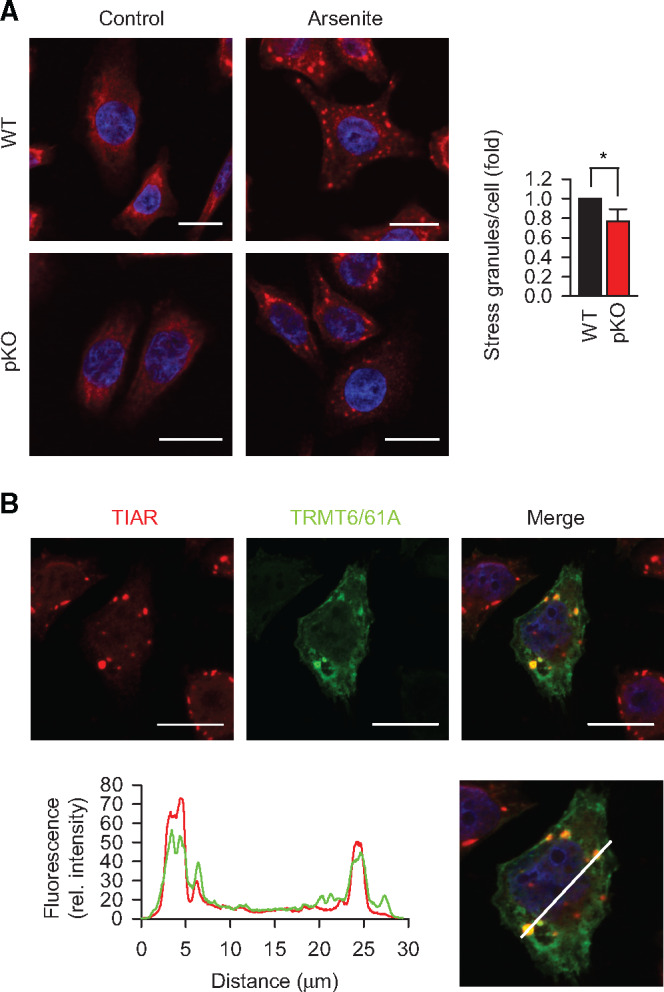
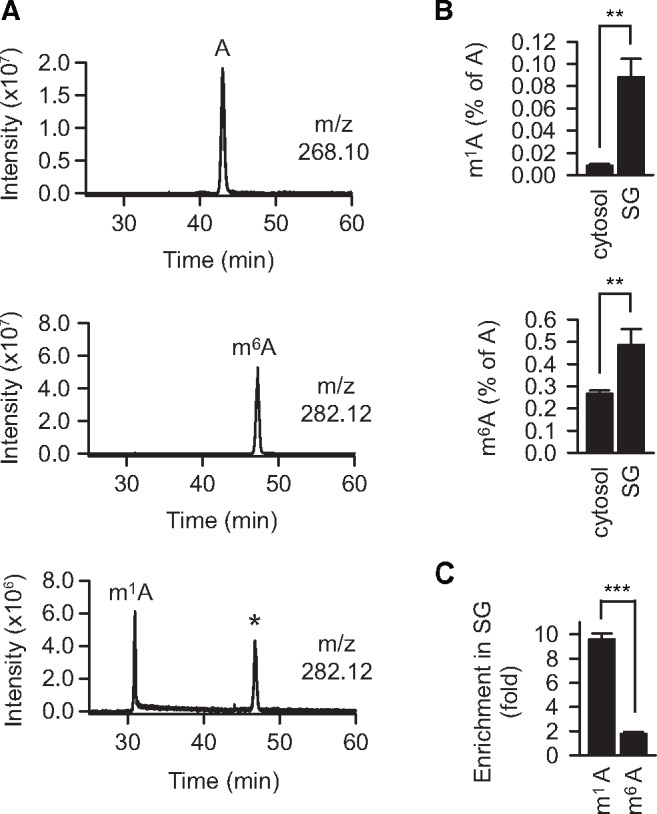
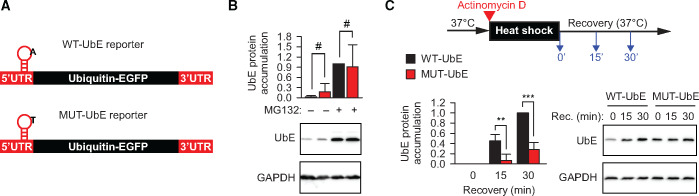
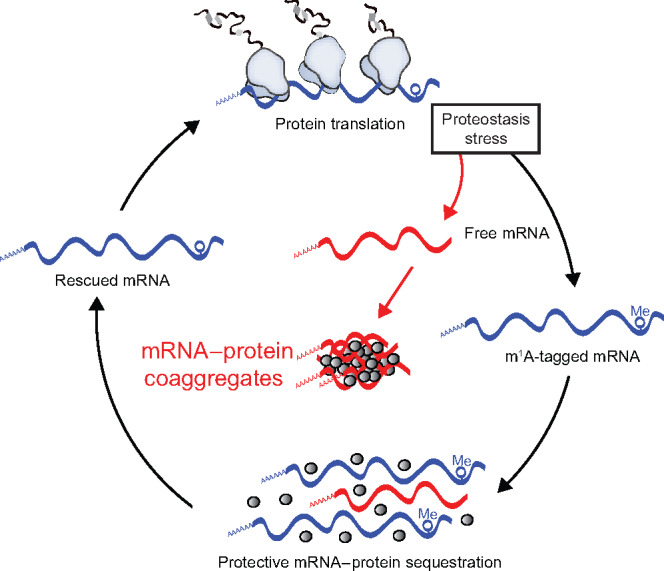
Post-transcriptional methylation of N6-adenine and N1-adenine can affect transcriptome turnover and translation. Furthermore, the regulatory function of N6-methyladenine (m6A) during heat shock has been uncovered, including the enhancement of the phase separation potential of RNAs. In response to acute stress, e.g. heat shock, the orderly sequestration of mRNAs in stress granules (SGs) is considered important to protect transcripts from the irreversible aggregation. Until recently, the role of N1-methyladenine (m1A) on mRNAs during acute stress response remains largely unknown. Here we show that the methyltransferase complex TRMT6/61A, which generates the m1A tag, is involved in transcriptome protection during heat shock. Our bioinformatics analysis indicates that occurrence of the m1A motif is increased in mRNAs known to be enriched in SGs. Accordingly, the m1A-generating methyltransferase TRMT6/61A accumulated in SGs and mass spectrometry confirmed enrichment of m1A in the SG RNAs. The insertion of a single methylation motif in the untranslated region of a reporter RNA leads to more efficient recovery of protein synthesis from that transcript after the return to normal temperature. Our results demonstrate far-reaching functional consequences of a minimal RNA modification on N1-adenine during acute proteostasis stress.
Keywords: N1-methyladenine; stress granules; stress response.
© The Author(s) (2020). Published by Oxford University Press on behalf of Journal of Molecular Cell Biology, IBCB, SIBS, CAS.
Figures
References
-
- Alriquet M., Martínez-Limón A., Hanspach G., et al. (2019). Assembly of proteins by free RNA during the early phase of proteostasis stress. J. Proteome Res. 18, 2835–284. - PubMed
-
- Blomen V.A., Májek P., Jae L.T., et al. (2015). Gene essentiality and synthetic lethality in haploid human cells. Science 350, 1092–1096. - PubMed
