

Improving Precise CRISPR Genome Editing by Small Molecules: Is there a Magic Potion?
- PMID: 32466303
- PMCID: PMC7291049
- DOI: 10.3390/cells9051318
Improving Precise CRISPR Genome Editing by Small Molecules: Is there a Magic Potion?
Abstract
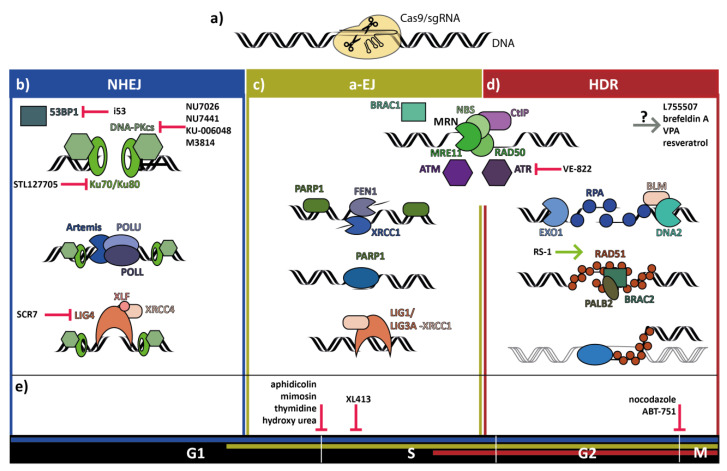
Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) genome editing has become a standard method in molecular biology, for the establishment of genetically modified cellular and animal models, for the identification and validation of drug targets in animals, and is heavily tested for use in gene therapy of humans. While the efficiency of CRISPR mediated gene targeting is much higher than of classical targeted mutagenesis, the efficiency of CRISPR genome editing to introduce defined changes into the genome is still low. Overcoming this problem will have a great impact on the use of CRISPR genome editing in academic and industrial research and the clinic. This review will present efforts to achieve this goal by small molecules, which modify the DNA repair mechanisms to facilitate the precise alteration of the genome.
Keywords: CRISPR efficiency; homology directed repair; low molecular weight compounds.
Conflict of interest statement
The authors declare the following competing interests: N.B. and C.B. declare no conflict of interest. M.M. is an employee and shareholder of AstraZeneca. S.W. is an industrial PhD student at AstraZeneca.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
