

Differential Complex Formation via Paralogs in the Human Sin3 Protein Interaction Network
- PMID: 32467258
- PMCID: PMC8143632
- DOI: 10.1074/mcp.RA120.002078
Differential Complex Formation via Paralogs in the Human Sin3 Protein Interaction Network
Abstract
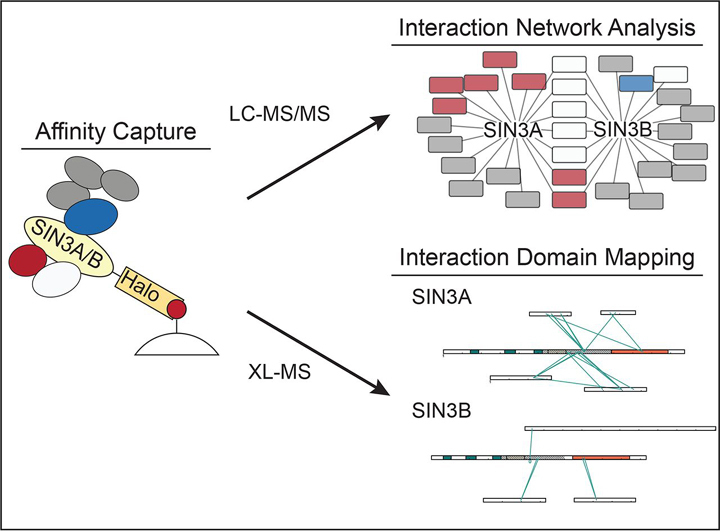
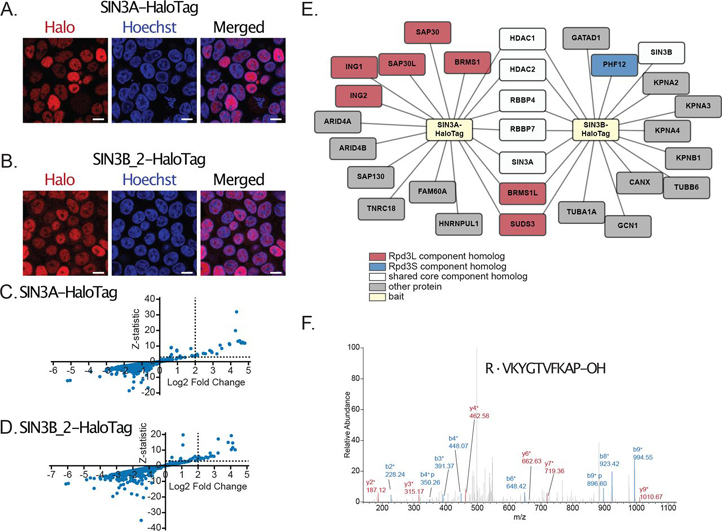
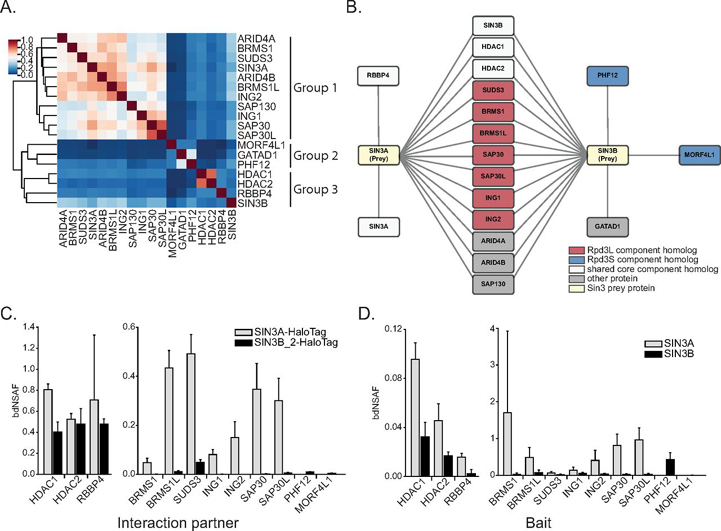
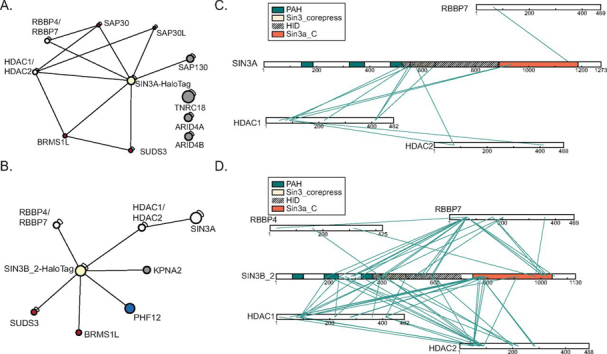
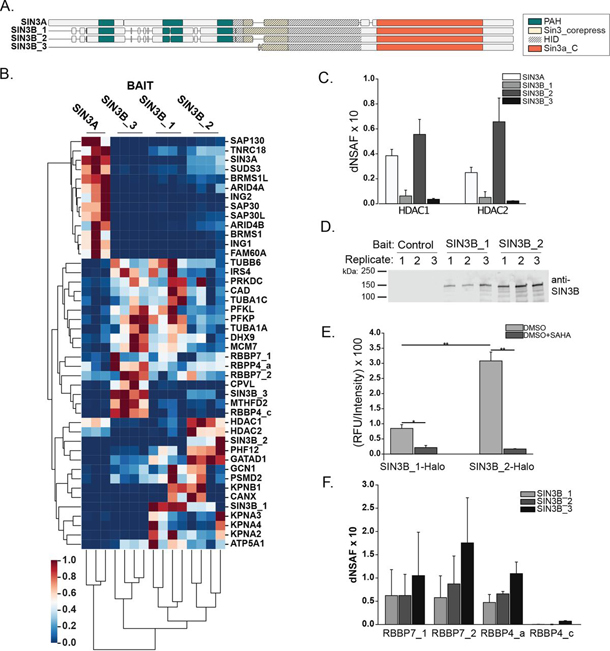
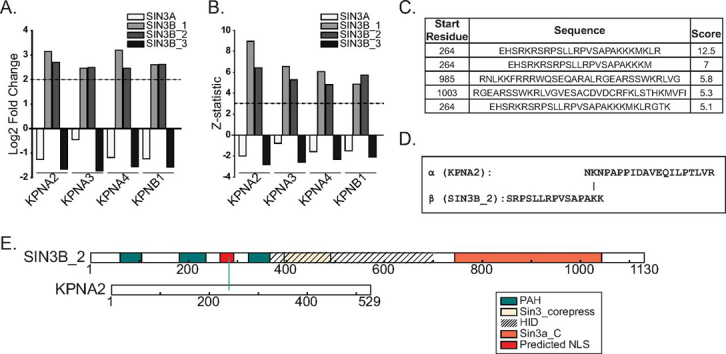
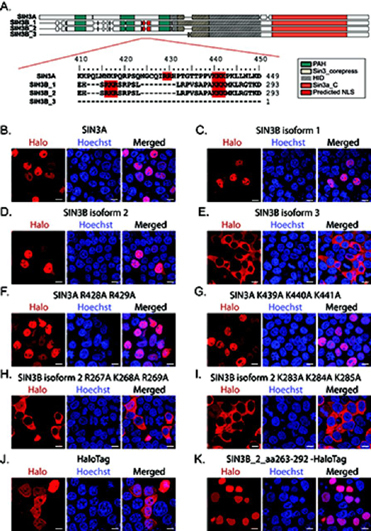
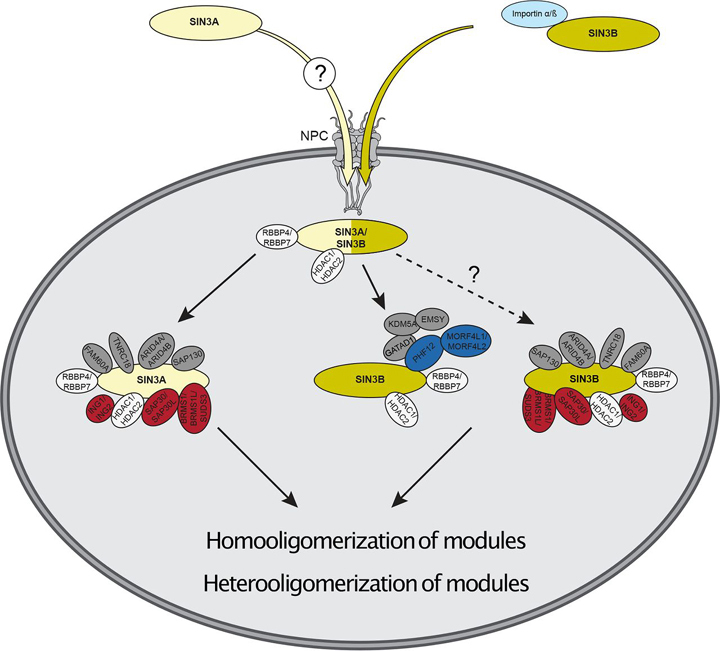
Despite the continued analysis of HDAC inhibitors in clinical trials, the heterogeneous nature of the protein complexes they target limits our understanding of the beneficial and off-target effects associated with their application. Among the many HDAC protein complexes found within the cell, Sin3 complexes are conserved from yeast to humans and likely play important roles as regulators of transcriptional activity. The presence of two Sin3 paralogs in humans, SIN3A and SIN3B, may result in a heterogeneous population of Sin3 complexes and contributes to our poor understanding of the functional attributes of these complexes. Here, we profile the interaction networks of SIN3A and SIN3B to gain insight into complex composition and organization. In accordance with existing data, we show that Sin3 paralog identity influences complex composition. Additionally, chemical cross-linking MS identifies domains that mediate interactions between Sin3 proteins and binding partners. The characterization of rare SIN3B proteoforms provides additional evidence for the existence of conserved and divergent elements within human Sin3 proteins. Together, these findings shed light on both the shared and divergent properties of human Sin3 proteins and highlight the heterogeneous nature of the complexes they organize.
Keywords: Chromatin function or biology; DSSO; SIN3; cross linking; epigenetics; histone deacetylase; nuclear translocation; pathway analysis; protein complex analysis; protein-protein interactions*; subcellular analysis; systems biology*.
© 2020 Adams et al.
Conflict of interest statement
Conflict of interest—Authors declare no competing interests.
Figures

References
-
- Link S., Spitzer R.M.M., Sana M., Torrado M., Völker-Albert M.C., Keilhauer E.C., Burgold T., Pünzeler S., Low J.K.K., Lindström I., Nist A., Regnard C., Stiewe T., Hendrich B., Imhof A., Mann M., Mackay J.P., Bartkuhn M., Hake S.B. PWWP2A binds distinct chromatin moieties and interacts with an MTA1-specific core NuRD complex. Nat. Commun. 2018;9:4300. - PMC - PubMed
-
- Carrozza M.J., Li B., Florens L., Suganuma T., Swanson S.K., Lee K.K., Shia W.J., Anderson S., Yates J., Washburn M.P., Workman J.L. Histone H3 methylation by Set2 directs deacetylation of coding regions by Rpd3S to suppress spurious intragenic transcription. Cell. 2005;123:581–592. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
