

Review
doi: 10.1038/s41576-020-0241-0.
Epub 2020 May 28.
Haematopoietic stem cell self-renewal in vivo and ex vivo
Affiliations
- PMID: 32467607
- PMCID: PMC7894993
- DOI: 10.1038/s41576-020-0241-0
Item in Clipboard
Review
Haematopoietic stem cell self-renewal in vivo and ex vivo
Nat Rev Genet.
2020 Sep.
Abstract
The self-renewal capacity of multipotent haematopoietic stem cells (HSCs) supports blood system homeostasis throughout life and underlies the curative capacity of clinical HSC transplantation therapies. However, despite extensive characterization of the HSC state in the adult bone marrow and embryonic fetal liver, the mechanism of HSC self-renewal has remained elusive. This Review presents our current understanding of HSC self-renewal in vivo and ex vivo, and discusses important advances in ex vivo HSC expansion that are providing new biological insights and offering new therapeutic opportunities.
Conflict of interest statement
Competing interests
H.N. is a co-founder and shareholder in Megakaryon and Century Therapeutics.
Figures
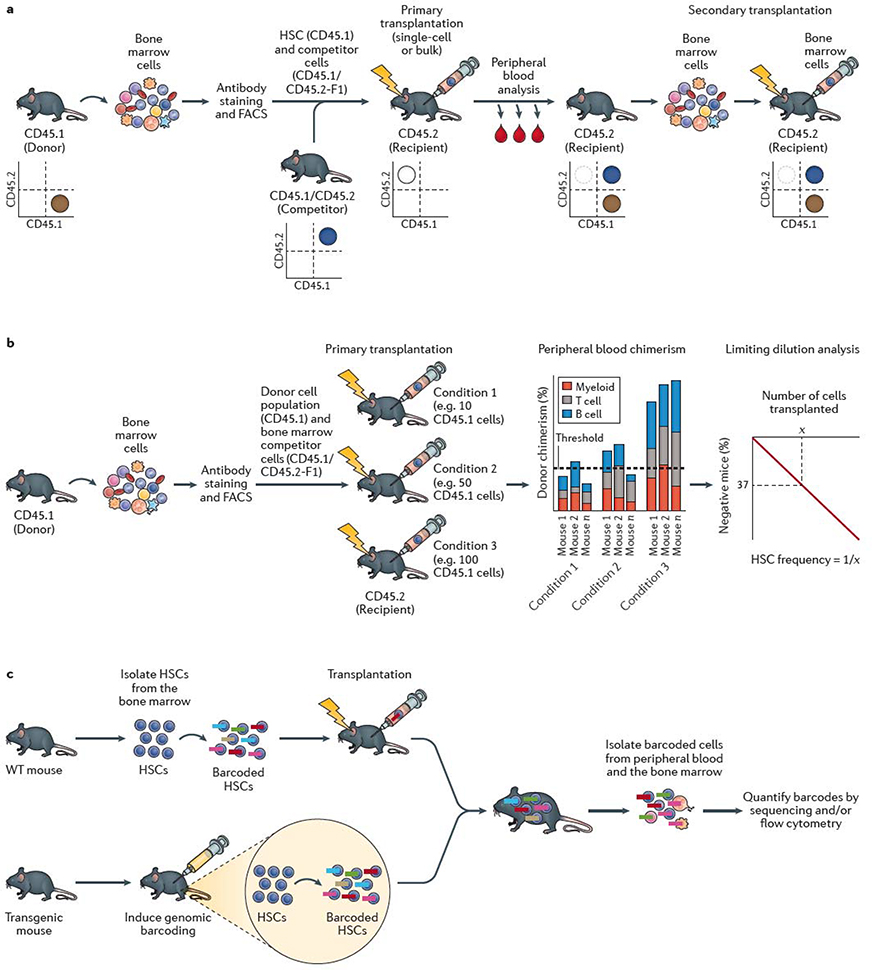
a | The standard mouse haematopoietic stem cell (HSC) transplantation assay uses the congenic CD45.1/CD45.2 system. Bone marrow cells are collected from a donor mouse bearing the CD45.1 allele from which HSCs are isolated using fluorescence-actiivated cell sorting (FACS). HSCs are mixed with competitor (also termed helper) bone marrow cells from CD45.1 × CD45.2 F1 mice and transplanted into a primary recipient mouse bearing the CD45.2 allele. Donor cell reconstitution kinetics can be determined within the peripheral blood and bone marrow using monoclonal antibodies and flow cytometric analysis. Flow cytometric quantification of donor chimerism within the peripheral blood is usually performed regularly over 16–24 weeks, and bone marrow analyzed at the study endpoint. As annotated in the representative flow cytometry plots, donor hematopoietic cells are CD45.1+, competitor hematopoietic cells are CD45.1+CD45.2+, while recipient hematopoietic cells are CD45.2+ (although irradation leads to loss of endogenous CD45.2+ haematopoietic cells in the transplant recipient). Bone marrow cells from the primary recipient mouse are transplanted into a secondary recipient mouse bearing the CD45.2 allele to confirm HSC self-renewal activity. Depending on the experimental design, CD45.1 recipient mice and CD45.2 donor mice may also be used. b | Limiting dilution analysis (LDA) can be used to estimate the frequency of HSCs within a cell population by transplantion of varying doses of cells into multiple mice. Presence or absence of an HSC(s) within the donor population is determined by assessing long-term peripheral blood reconstitution and LDA performed based on the number of positive versus negative mice as a function of the donor cell dose. Various reconstitution thresholds have been used in the field, but it is typically set at 1% donor peripheral blood chimerism. Based on Poisson statistics, the number of cells (X) that result in 37% negative mice is equal to the 1/HSC frequency. c | HSC self-renewal can be tracked using barcoding (or other genetic labelling) technologies. Barcoding can be performed ex vivo, by isolating HSCs from the bone marrow and labeling them in vitro with genetic barcodes (which can be applied using various methods, including transduction with a lentiviral library) and transplanted into recipient mice. Barcoding can also be performed directly in vivo, usually via inducing activity of a recombinase-based or transposon-based ‘shuffling’ of genetic sequences to generate unique genomic sequences in each HSC. The output of individual HSCs can be determined by quantifying the barcode abundance within peripheral blood and/or bone marrow cell populations. In vivo lineage tracing technologies can be used to track HSC self-renewal in the native bone marrow without the requirement for transplantation.
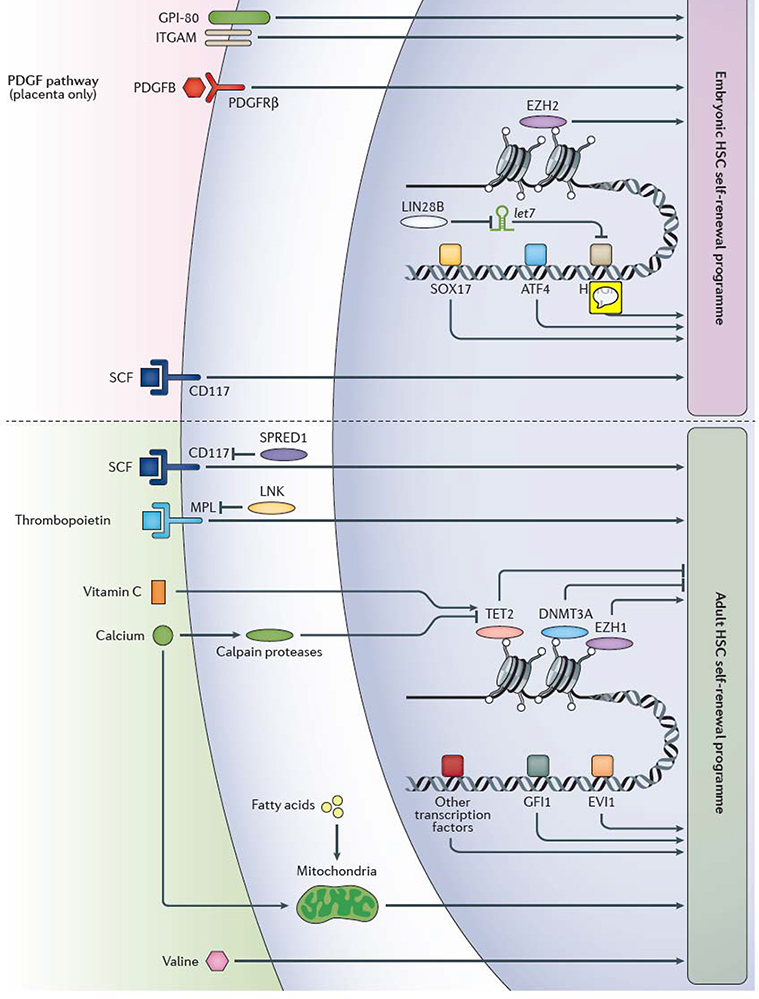
Various extrinsic and intrinsic regulators of embryonic and adult haematopoietic stem cell (HSC) self-renewal have been identified in mice and humans, however, our knowledge of the mechanism of HSC self-renewal remain incomplete. Factors that regulate HSC self-renewal in adult mice have been primarily studied in bone marrow HSCs, whereas factors that regulate HSC self-renewal in the mouse embryo have been mainly studied in fetal liver HSCs, except the platelet-derived growth factor (PDGF) pathway, which has been shown to regulate HSC differentiation only in the placenta. Our knowledge of the exact relationships between the various self-renewal factors is often lacking, although protein lin28 homolog B (Lin28b), the microRNA let-7 and high mobility group protein HMGIC (Hmga2) are known to participate in the same embryonic HSC self-renewal pathway. Multiple transcription factors including Sox17 and Atf4, as well as epigenetic regulators such as the polycomb repressive complex 2 component histone-lysine N-methyltransferase Ezh2, influence gene expression to promote HSC self-renewal in the mouse embryo. HSC self-renewal in adulthood is also regulated within the nucleus, including by the epigenetic regulatory molecules methylcytosine dioxygenase Tet2, DNA (cytosine-5)-methyltransferase 3A (Dnmt3a), the polycomb repressive complex 1 component Ezh1, and transcription factors including Gfi1 and Evi1, amongst others. CD117, Scf receptor Kit; Evi1, Ectropic virus integration site protein 1 homolog, histone-lysine N-methyltransferase MECOM; GPI-80, vanin 2; ITGAM, integrin αM; Lnk, SH2B adapter protein 3; Mpl, myeloproliferative leukemia protein, also known as thrombopoietin receptor; Pdgfb, platelet-derived growth factor subunit B; Pdgfrβ, platelet-derived growth factor receptor β; Scf, stem cell factor (also known as Kit ligand); Spred1, sprouty-related, EVH1-domain-containing protein 1; transcription factors, TFs.
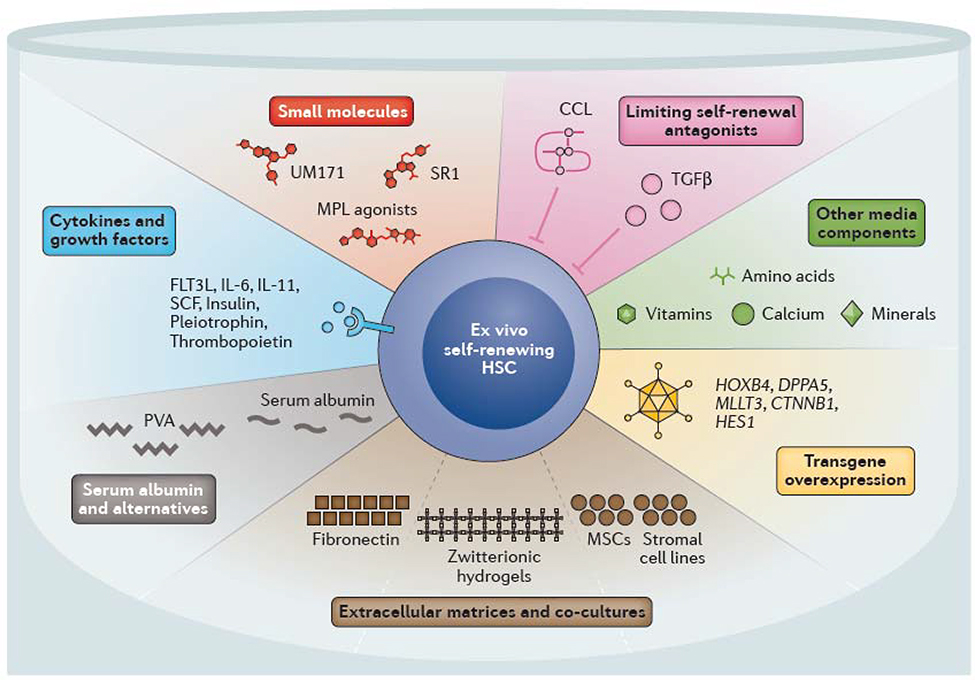
Multiple strategies have been developed to support ex vivo haematopoietic stem cell (HSC) self-renewal. Many of the cytokines and growth factors that have been found to regulate in vivo self-renewal have been used to promote ex vivo self-renewal, including thrombopoietin and stem cell factor (SCF, also known as Kit ligand). Multiple small molecules and agonists have been identified that mimic or promote self-renewal in combination with these cytokines and growth factors. Specifically, StemRegenin 1 (SR1) and UM171 promote expansion of human cord blood HSCs. Other strategies include preventing the accumulation of endogenous self-renewal inhibitors, including transforming growth factor β (TGFβ) and C-C motif chemokine ligands (CCLs) by batch fed culture methods or complete media changes. Changes in other media components also influence ex vivo HSC stability (for example, low calcium levels promote HSC maintenance). Serum albumin has long been used in HSC culture and expansion systems, although it can can be replaced with polyvinyl alcohol (PVA). Extracellular matrices consisting of fibronectin or hydrogels and/or co-culture with mesenchymal stem cells (MSCs) or stromal cells can also help to promote HSC self-renewal. Genetic perturbations, including overexpression of transcription factors such as MTTL3 (AF9), homeobox protein Hoxb4 and developmental pluripotency-associated protein 5A (Dppa5) as well as upregulation of signaling pathways (Notch and Wnt) also promote ex vivo HSC self-renewal. CD117, Scf receptor Kit; FLT3L, FMS-like tyrosine kinase 3 ligand; IL, interleukin; MPL, myeloproliferative leukemia protein, also known as thrombopoietin receptor.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
