

The Potency of an Anti-MERS Coronavirus Subunit Vaccine Depends on a Unique Combinatorial Adjuvant Formulation
- PMID: 32471056
- PMCID: PMC7350031
- DOI: 10.3390/vaccines8020251
The Potency of an Anti-MERS Coronavirus Subunit Vaccine Depends on a Unique Combinatorial Adjuvant Formulation
Abstract
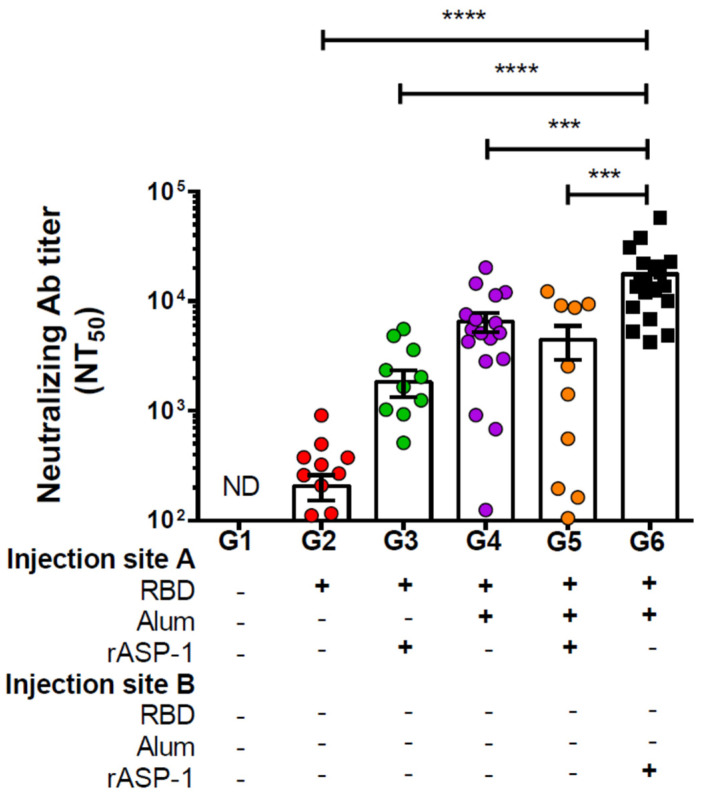
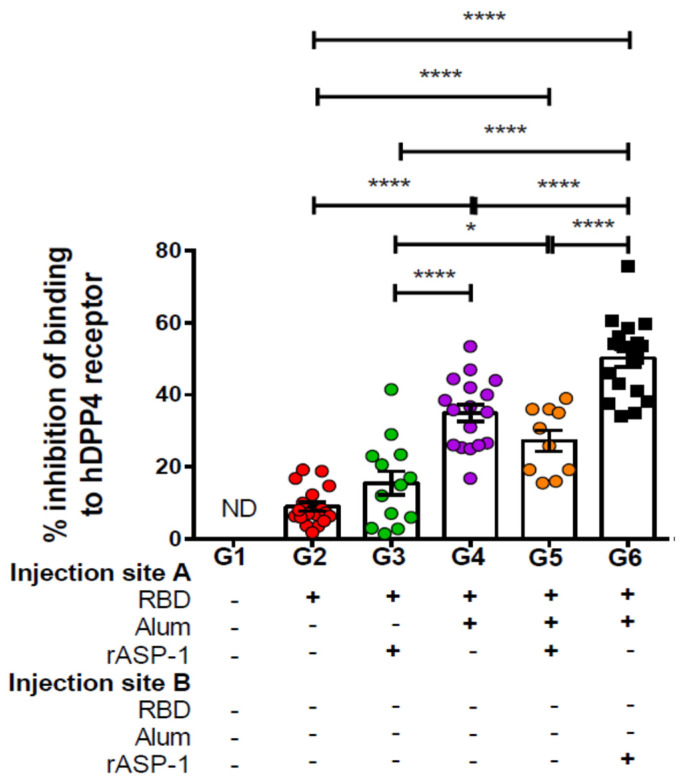
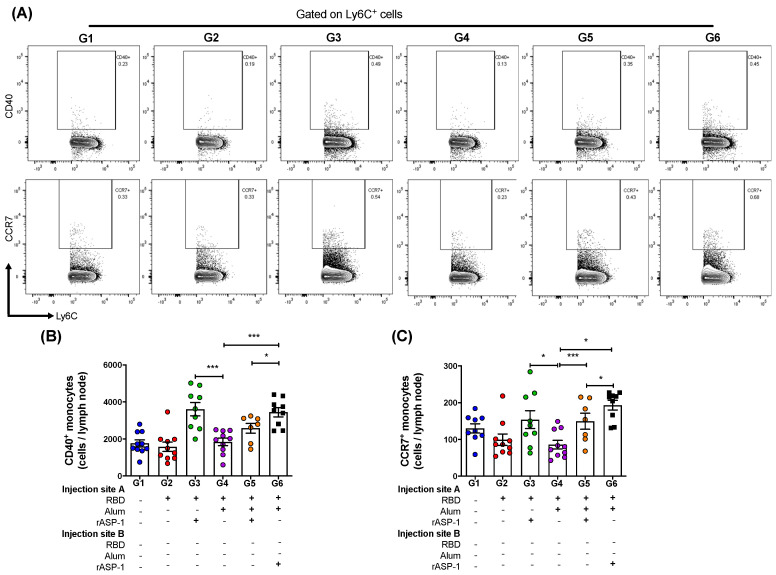
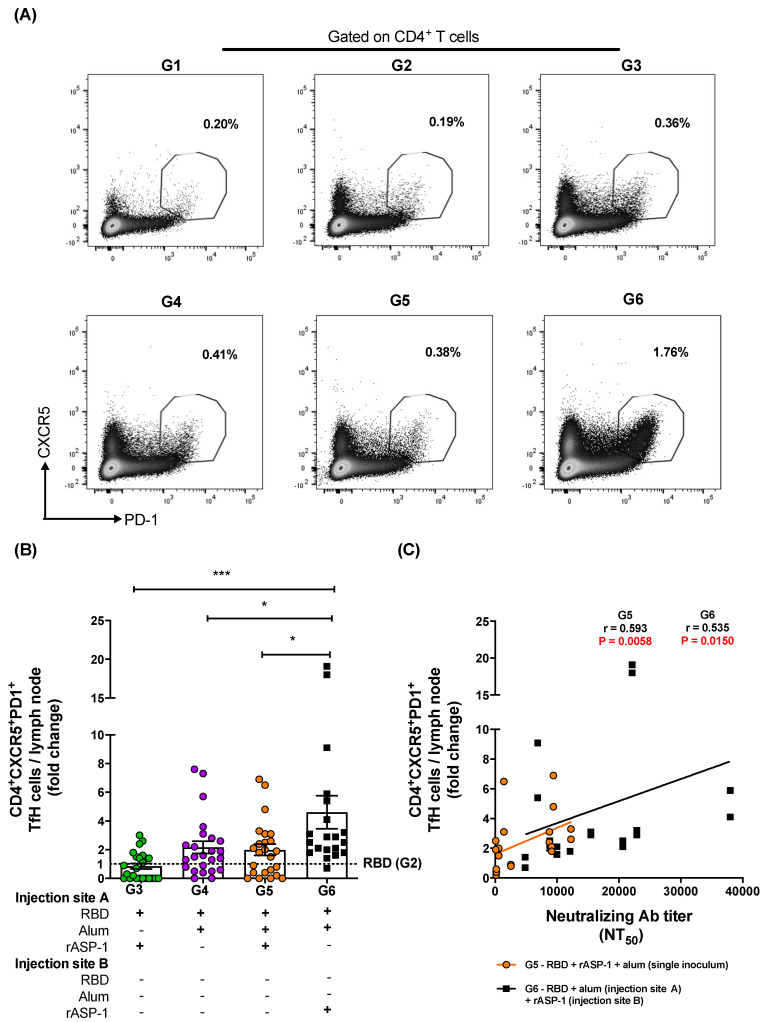
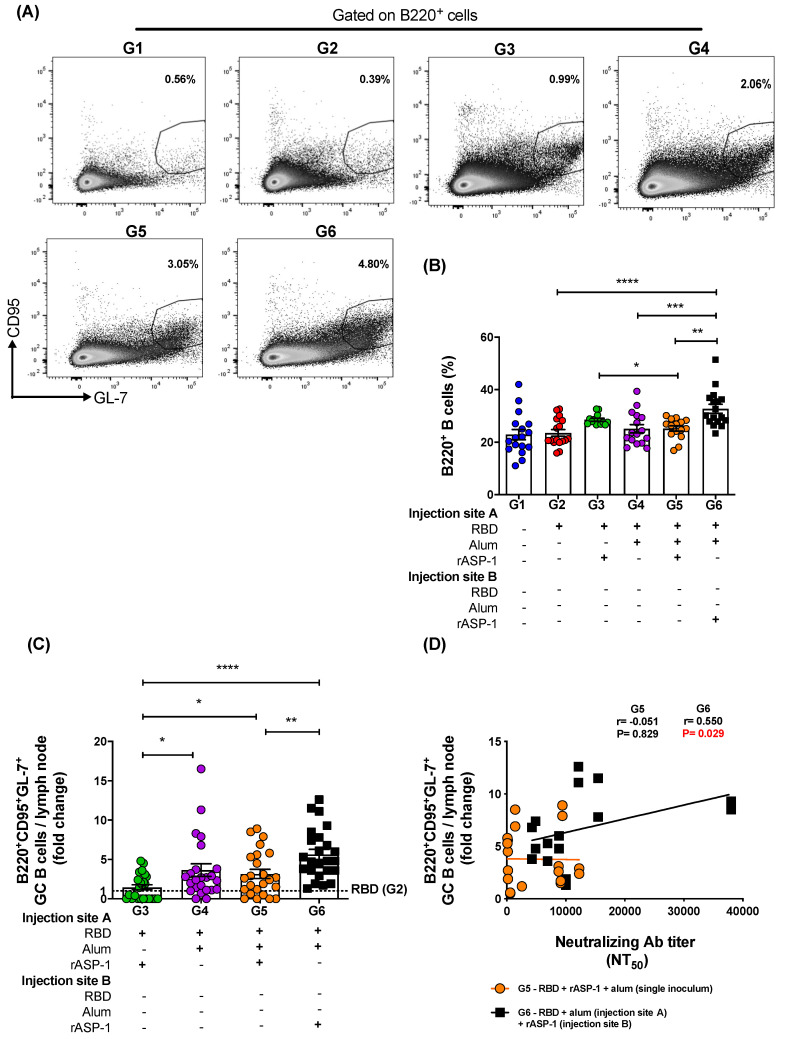
Vaccination is one of the most successful strategies to prevent human infectious diseases. Combinatorial adjuvants have gained increasing interest as they can stimulate multiple immune pathways and enhance the vaccine efficacy of subunit vaccines. We investigated the adjuvanticity of Aluminum (alum) in combination with rASP-1, a protein adjuvant, using the Middle East respiratory syndrome coronavirus MERS-CoV receptor-binding-domain (RBD) vaccine antigen. A highly enhanced anti-MERS-CoV neutralizing antibody response was induced when mice were immunized with rASP-1 and the alum-adjuvanted RBD vaccine in two separate injection sites as compared to mice immunized with RBD + rASP-1 + alum formulated into a single inoculum. The antibodies produced also significantly inhibited the binding of RBD to its cell-associated receptor. Moreover, immunization with rASP-1 co-administered with the alum-adjuvanted RBD vaccine in separate sites resulted in an enhanced frequency of TfH and GC B cells within the draining lymph nodes, both of which were positively associated with the titers of the neutralizing antibody response related to anti-MERS-CoV protective immunity. Our findings not only indicate that this unique combinatorial adjuvanted RBD vaccine regimen improved the immunogenicity of RBD, but also point to the importance of utilizing combinatorial adjuvants for the induction of synergistic protective immune responses.
Keywords: MERS-CoV; T follicular helper cells; adjuvant combination; adjuvants; aluminum; functional antibody responses; germinal center B cells; rASP-1; receptor-binding domain; synergy.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
