

TRIM72 promotes alveolar epithelial cell membrane repair and ameliorates lung fibrosis
- PMID: 32471489
- PMCID: PMC7257505
- DOI: 10.1186/s12931-020-01384-2
TRIM72 promotes alveolar epithelial cell membrane repair and ameliorates lung fibrosis
Abstract
Background: Chronic tissue injury was shown to induce progressive scarring in fibrotic diseases such as idiopathic pulmonary fibrosis (IPF), while an array of repair/regeneration and stress responses come to equilibrium to determine the outcome of injury at the organ level. In the lung, type I alveolar epithelial (ATI) cells constitute the epithelial barrier, while type II alveolar epithelial (ATII) cells play a pivotal role in regenerating the injured distal lungs. It had been demonstrated that eukaryotic cells possess repair machinery that can quickly patch the damaged plasma membrane after injury, and our previous studies discovered the membrane-mending role of Tripartite motif containing 72 (TRIM72) that expresses in a limited number of tissues including the lung. Nevertheless, the role of alveolar epithelial cell (AEC) repair in the pathogenesis of IPF has not been examined yet.
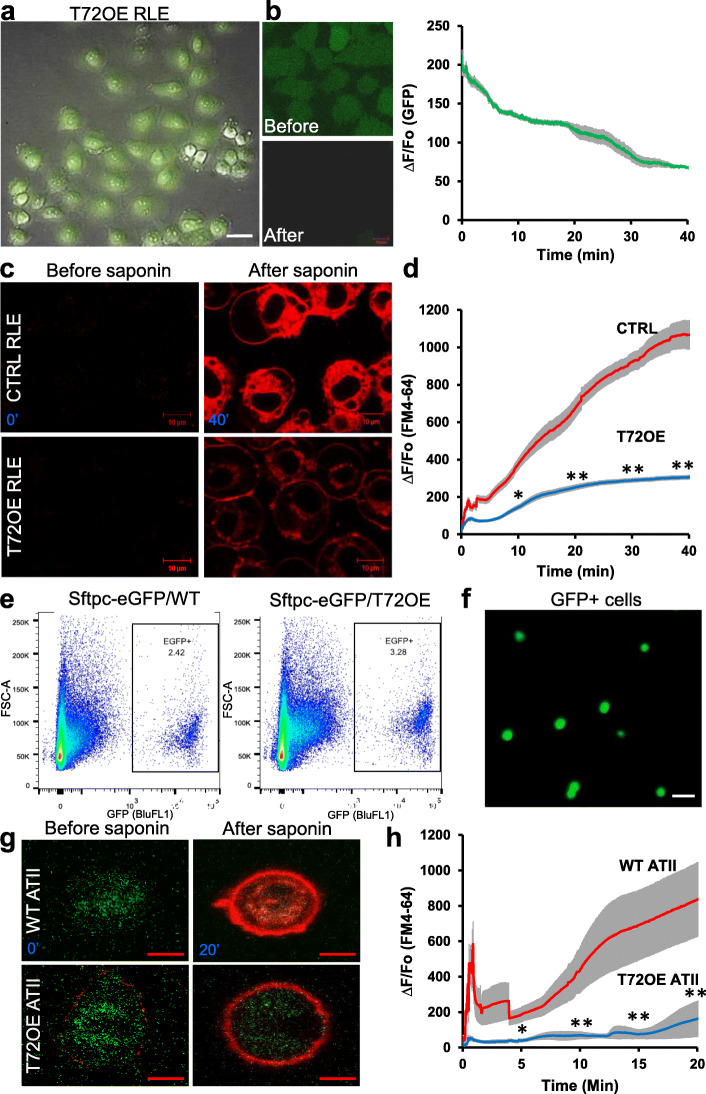
Method: In this study, we tested the specific roles of TRIM72 in the repair of ATII cells and the development of lung fibrosis. The role of membrane repair was accessed by saponin assay on isolated primary ATII cells and rat ATII cell line. The anti-fibrotic potential of TRIM72 was tested with bleomycin-treated transgenic mice.
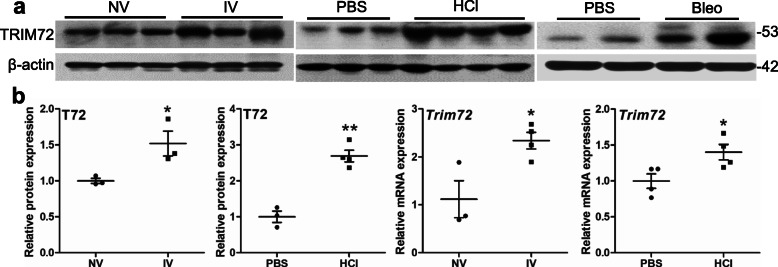
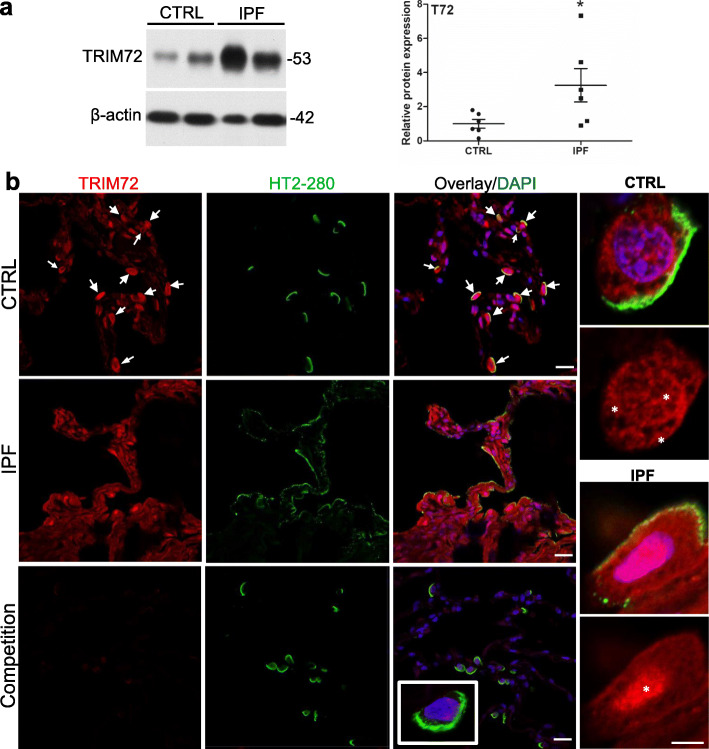
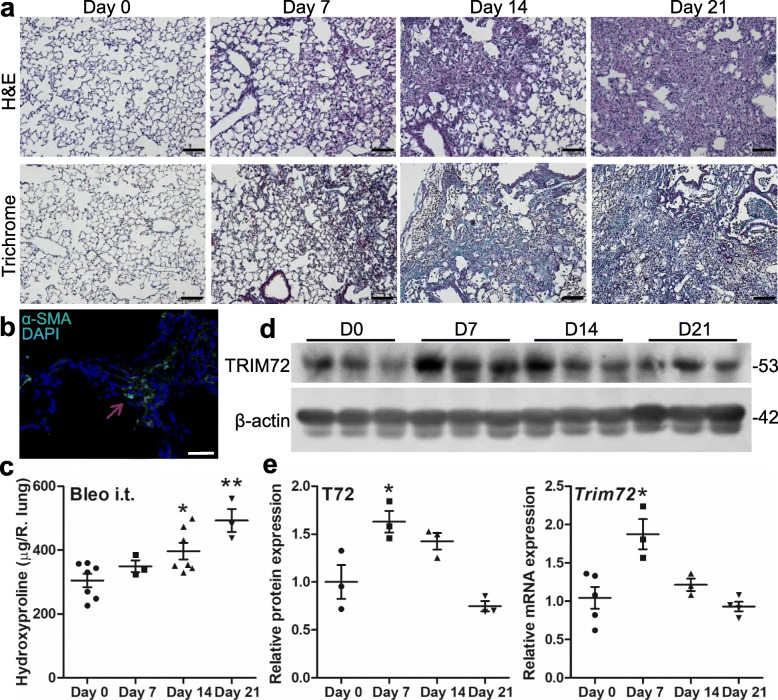
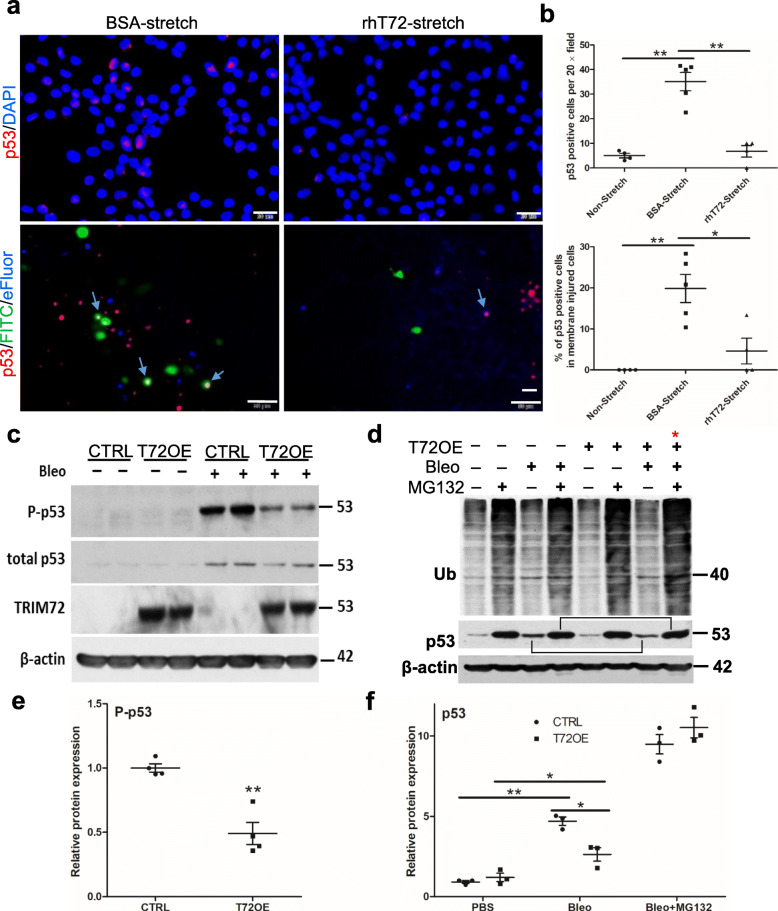
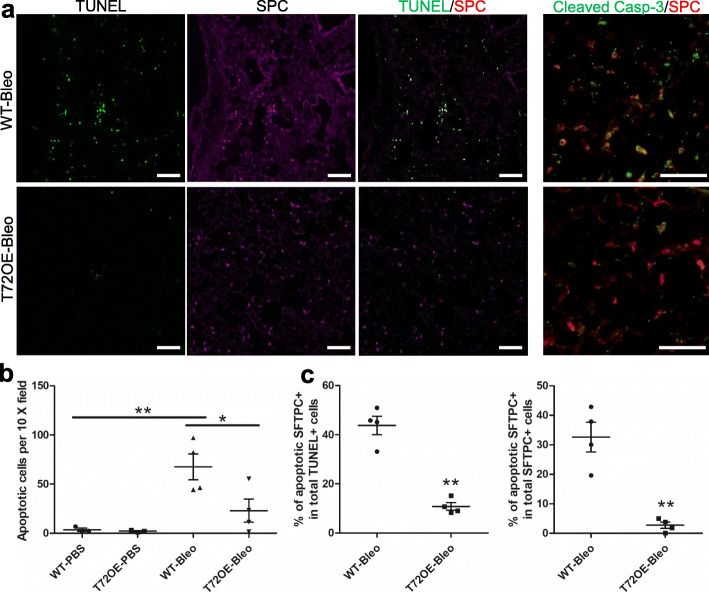
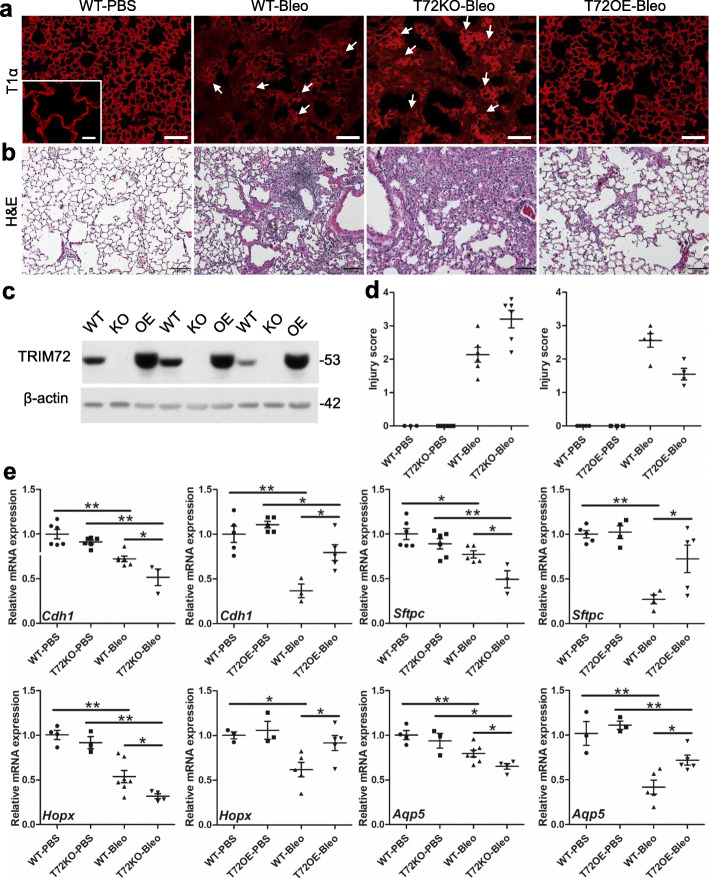
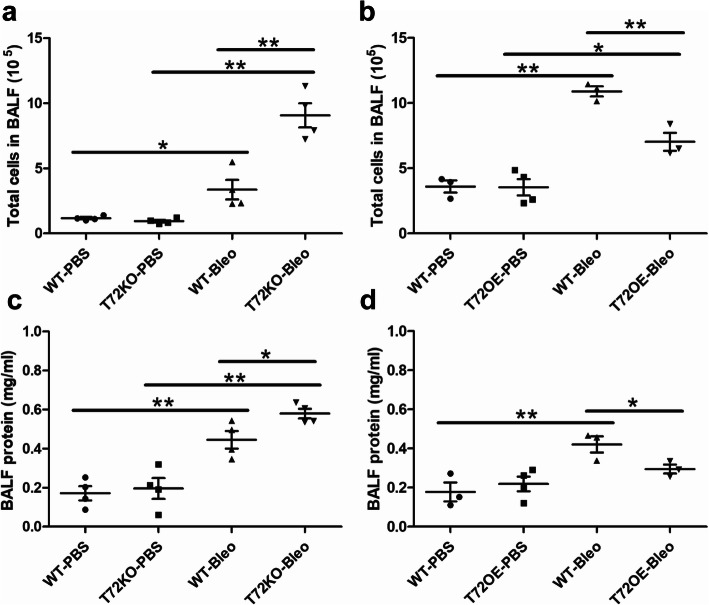
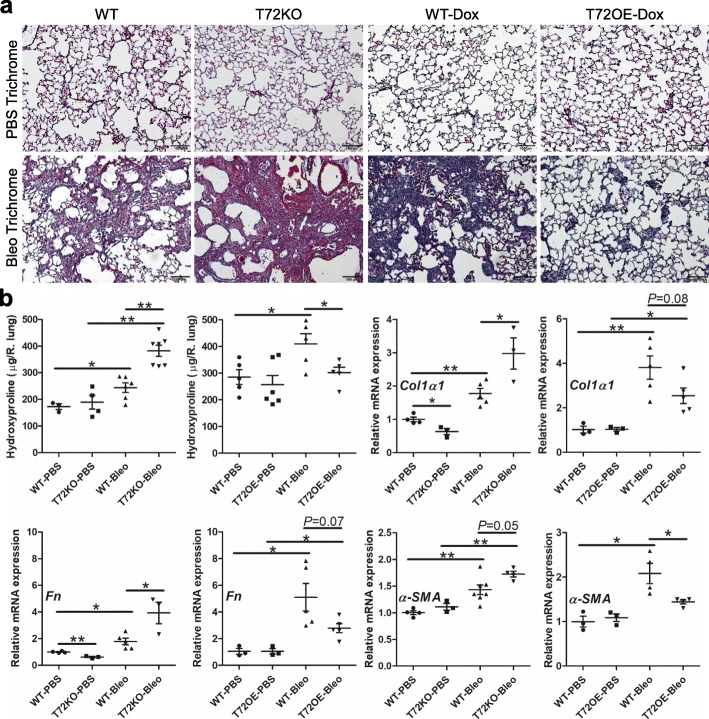
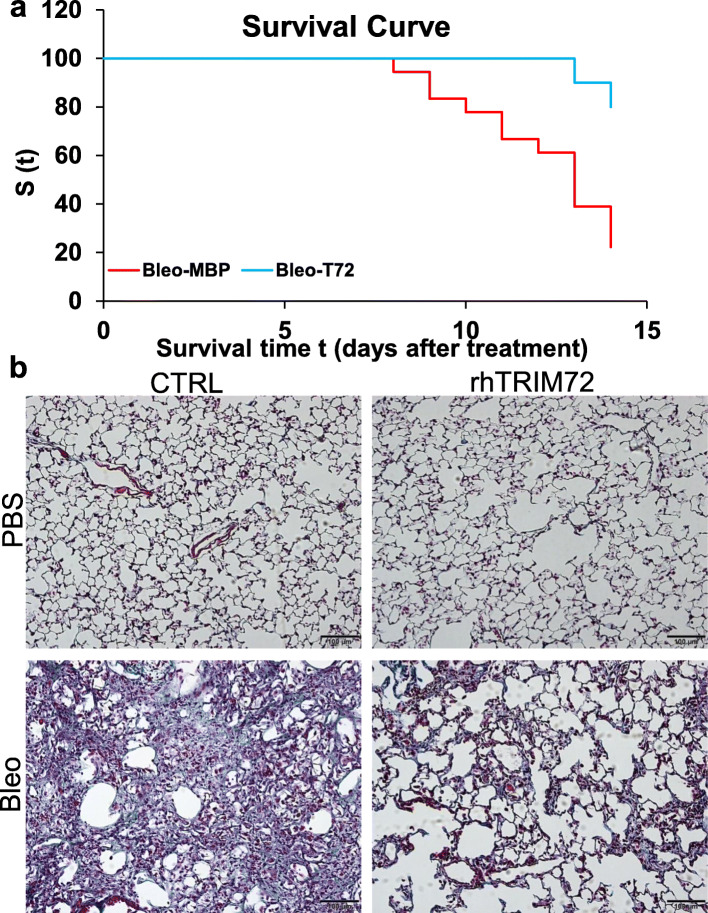
Results: We showed that TRIM72 was upregulated following various injuries and in human IPF lungs. However, TRIM72 expression in ATII cells of the IPF lungs had aberrant subcellular localization. In vitro studies showed that TRIM72 repairs membrane injury of immortalized and primary ATIIs, leading to inhibition of stress-induced p53 activation and reduction in cell apoptosis. In vivo studies demonstrated that TRIM72 protects the integrity of the alveolar epithelial layer and reduces lung fibrosis.
Conclusion: Our results suggest that TRIM72 protects injured lungs and ameliorates fibrosis through promoting post-injury repair of AECs.
Keywords: Alveolar epithelial cells; Apoptosis; Idiopathic pulmonary fibrosis; Membrane repair; Tripartite motif family protein 72.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Dynamic expression of HOPX in alveolar epithelial cells reflects injury and repair during the progression of pulmonary fibrosis.Sci Rep. 2018 Aug 28;8(1):12983. doi: 10.1038/s41598-018-31214-x. Sci Rep. 2018. PMID: 30154568 Free PMC article.
-
Serpine 1 induces alveolar type II cell senescence through activating p53-p21-Rb pathway in fibrotic lung disease.Aging Cell. 2017 Oct;16(5):1114-1124. doi: 10.1111/acel.12643. Epub 2017 Jul 19. Aging Cell. 2017. PMID: 28722352 Free PMC article.
-
Chronic WNT/β-catenin signaling induces cellular senescence in lung epithelial cells.Cell Signal. 2020 Jun;70:109588. doi: 10.1016/j.cellsig.2020.109588. Epub 2020 Feb 26. Cell Signal. 2020. PMID: 32109549 Free PMC article.
-
Regeneration or Repair? The Role of Alveolar Epithelial Cells in the Pathogenesis of Idiopathic Pulmonary Fibrosis (IPF).Cells. 2022 Jun 30;11(13):2095. doi: 10.3390/cells11132095. Cells. 2022. PMID: 35805179 Free PMC article. Review.
-
Targeting Alveolar Repair in Idiopathic Pulmonary Fibrosis.Am J Respir Cell Mol Biol. 2021 Oct;65(4):347-365. doi: 10.1165/rcmb.2020-0476TR. Am J Respir Cell Mol Biol. 2021. PMID: 34129811 Free PMC article. Review.
Cited by
-
Plasma membrane integrity: implications for health and disease.BMC Biol. 2021 Apr 13;19(1):71. doi: 10.1186/s12915-021-00972-y. BMC Biol. 2021. PMID: 33849525 Free PMC article. Review.
-
Plasma membrane disruption (PMD) formation and repair in mechanosensitive tissues.Bone. 2021 Aug;149:115970. doi: 10.1016/j.bone.2021.115970. Epub 2021 Apr 21. Bone. 2021. PMID: 33892174 Free PMC article. Review.
-
Enhancing Membrane Repair Using Recombinant MG53/TRIM72 (rhMG53) Reduces Neurotoxicity in Alzheimer's Disease Models.Biomolecules. 2025 Mar 15;15(3):418. doi: 10.3390/biom15030418. Biomolecules. 2025. PMID: 40149954 Free PMC article.
-
Identification and Validation of Aging-Related Genes in Idiopathic Pulmonary Fibrosis.Front Genet. 2022 Feb 8;13:780010. doi: 10.3389/fgene.2022.780010. eCollection 2022. Front Genet. 2022. PMID: 35211155 Free PMC article.
-
Curculigoside Attenuates Endoplasmic Reticulum Stress-Induced Epithelial Cell and Fibroblast Senescence by Regulating the SIRT1-P300 Signaling Pathway.Antioxidants (Basel). 2024 Mar 29;13(4):420. doi: 10.3390/antiox13040420. Antioxidants (Basel). 2024. PMID: 38671868 Free PMC article.
References
-
- McNeil PL, Steinhardt RA. Plasma membrane disruption: repair, prevention, adaptation. Annu Rev Cell Dev Biol. 2003;19:697–731. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
