

Developmental regulation of canonical and small ORF translation from mRNAs
- PMID: 32471506
- PMCID: PMC7260771
- DOI: 10.1186/s13059-020-02011-5
Developmental regulation of canonical and small ORF translation from mRNAs
Abstract
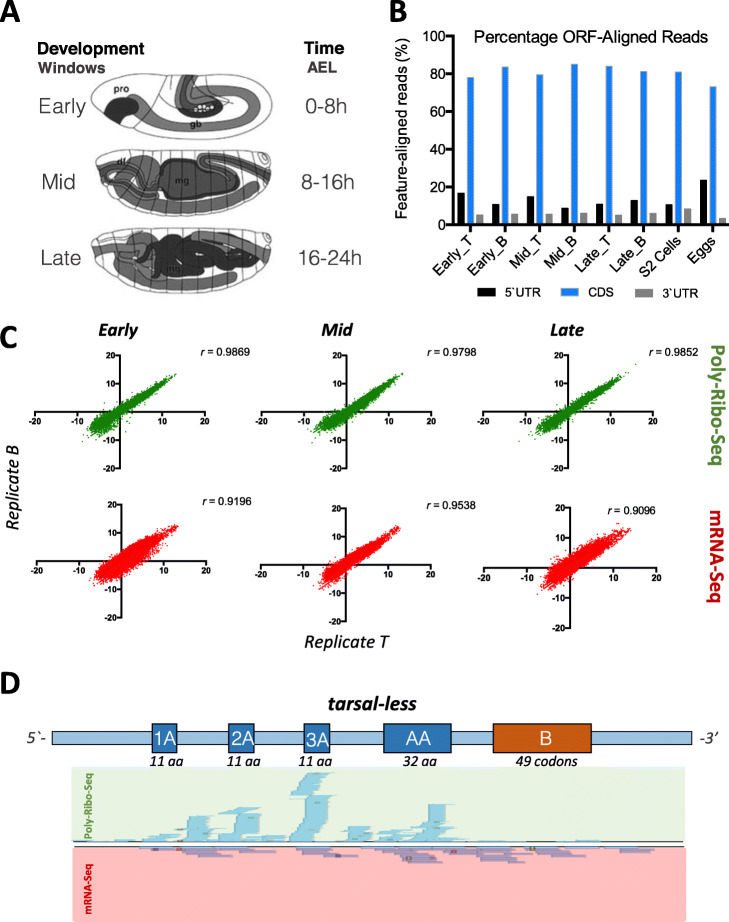
Background: Ribosomal profiling has revealed the translation of thousands of sequences outside annotated protein-coding genes, including small open reading frames of less than 100 codons, and the translational regulation of many genes. Here we present an improved version of Poly-Ribo-Seq and apply it to Drosophila melanogaster embryos to extend the catalog of in vivo translated small ORFs, and to reveal the translational regulation of both small and canonical ORFs from mRNAs across embryogenesis.
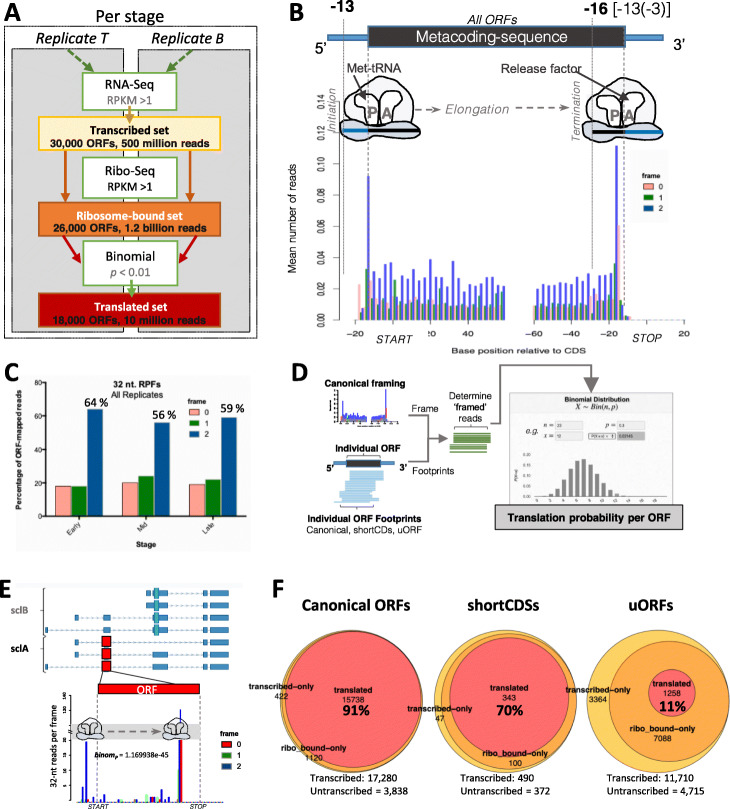
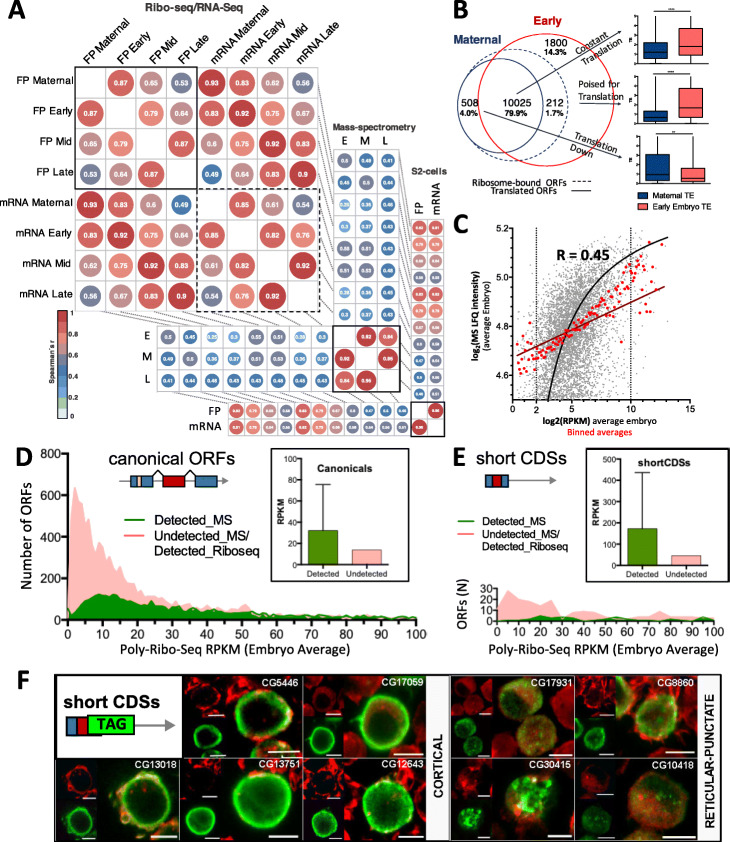
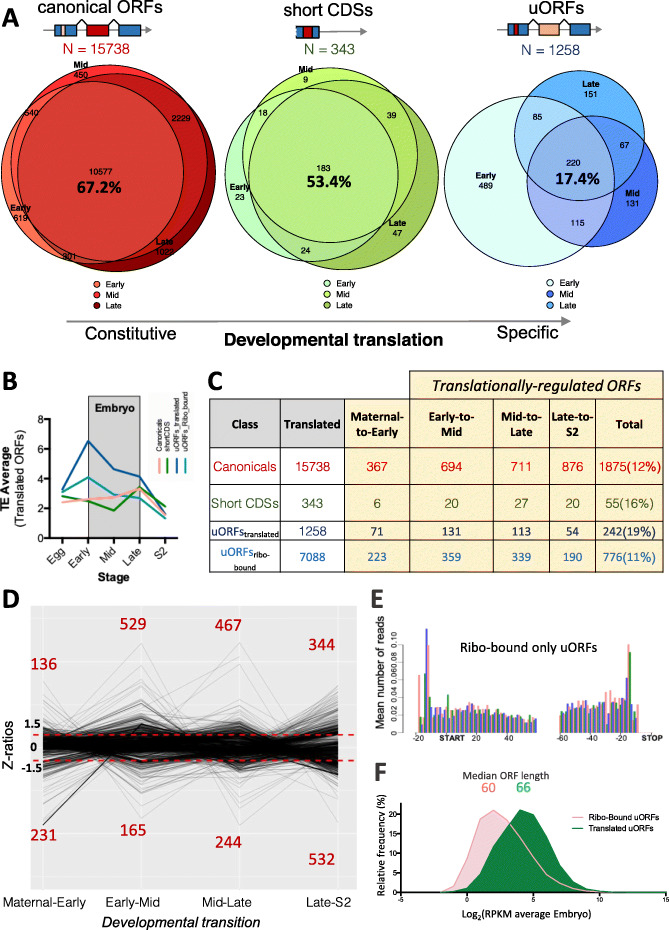
Results: We obtain highly correlated samples across five embryonic stages, with nearly 500 million putative ribosomal footprints mapped to mRNAs, and compare them to existing Ribo-Seq and proteomic data. Our analysis reveals, for the first time in Drosophila, footprints mapping to codons in a phased pattern, the hallmark of productive translation. We propose a simple binomial probability metric to ascertain translation probability. Our results also reveal reproducible ribosomal binding apparently not resulting in productive translation. This non-productive ribosomal binding seems to be especially prevalent amongst upstream short ORFs located in the 5' mRNA leaders, and amongst canonical ORFs during the activation of the zygotic translatome at the maternal-to zygotic transition.
Conclusions: We suggest that this non-productive ribosomal binding might be due to cis-regulatory ribosomal binding and to defective ribosomal scanning of ORFs outside periods of productive translation. Our results are compatible with the main function of upstream short ORFs being to buffer the translation of canonical canonical ORFs; and show that, in general, small ORFs in mRNAs display markers compatible with an evolutionary transitory state towards full coding function.
Keywords: Maternal to zygotic transition; Non-canonical translation; Poly-Ribo-Seq; Regulation of translation; Ribosomal binding; Ribosomal profiling; sORFs; smORFs; uORFs.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Calviello L, Mukherjee N, Wyler E, Zauber H, Hirsekorn A, Selbach M, Landthaler M, Obermayer B, Ohler U. Detecting actively translated open reading frames in ribosome profiling data. Nat Methods. 2016;13:165–170. - PubMed
-
- Mumtaz MA, Couso JP. Ribosomal profiling adds new coding sequences to the proteome. Biochem Soc Trans. 2015;43:1271–1276. - PubMed
