

A chemical inhibitor of heat shock protein 78 (HSP78) from Leishmania donovani represents a potential antileishmanial drug candidate
- PMID: 32471865
- PMCID: PMC7380179
- DOI: 10.1074/jbc.RA120.014587
A chemical inhibitor of heat shock protein 78 (HSP78) from Leishmania donovani represents a potential antileishmanial drug candidate
Abstract
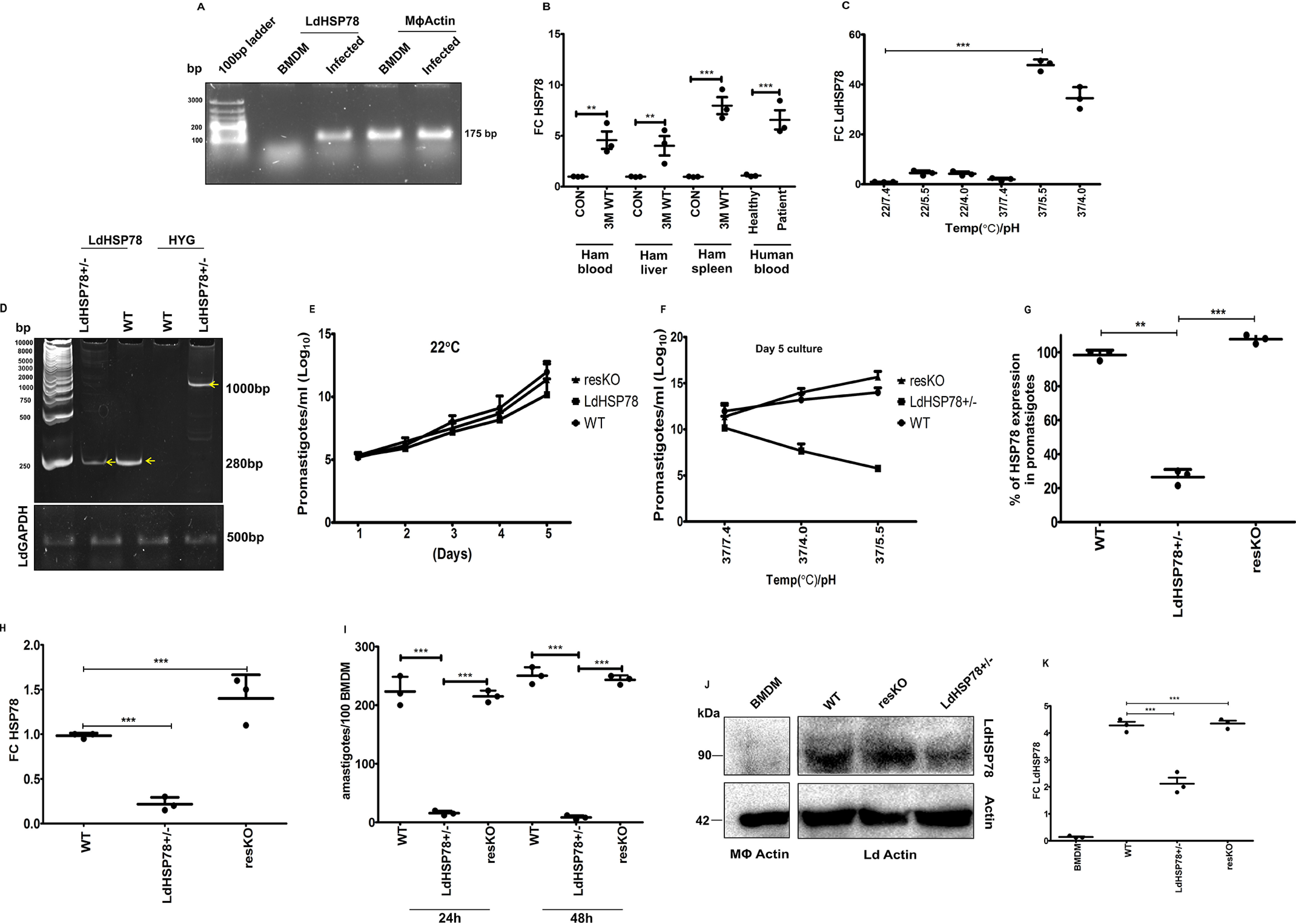
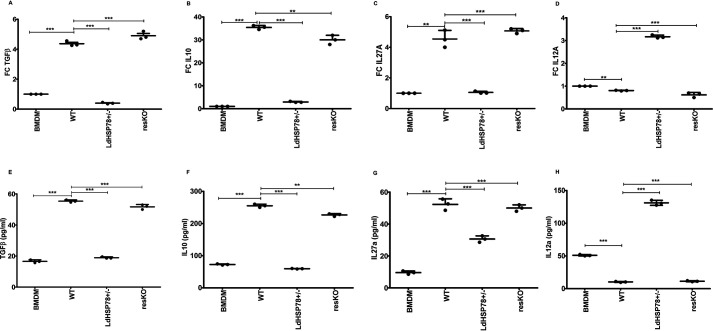
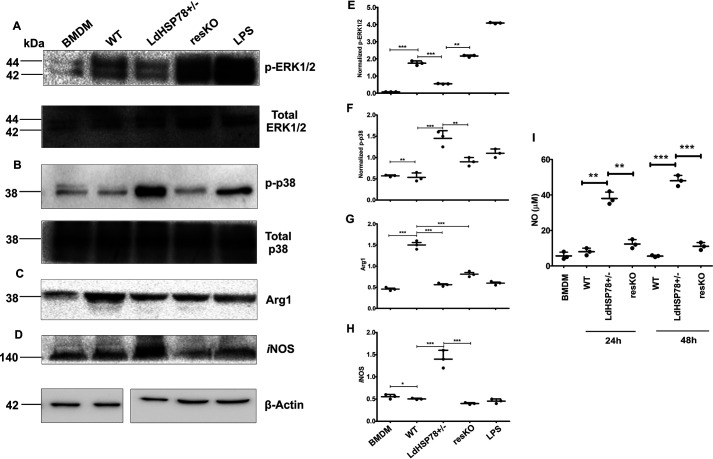
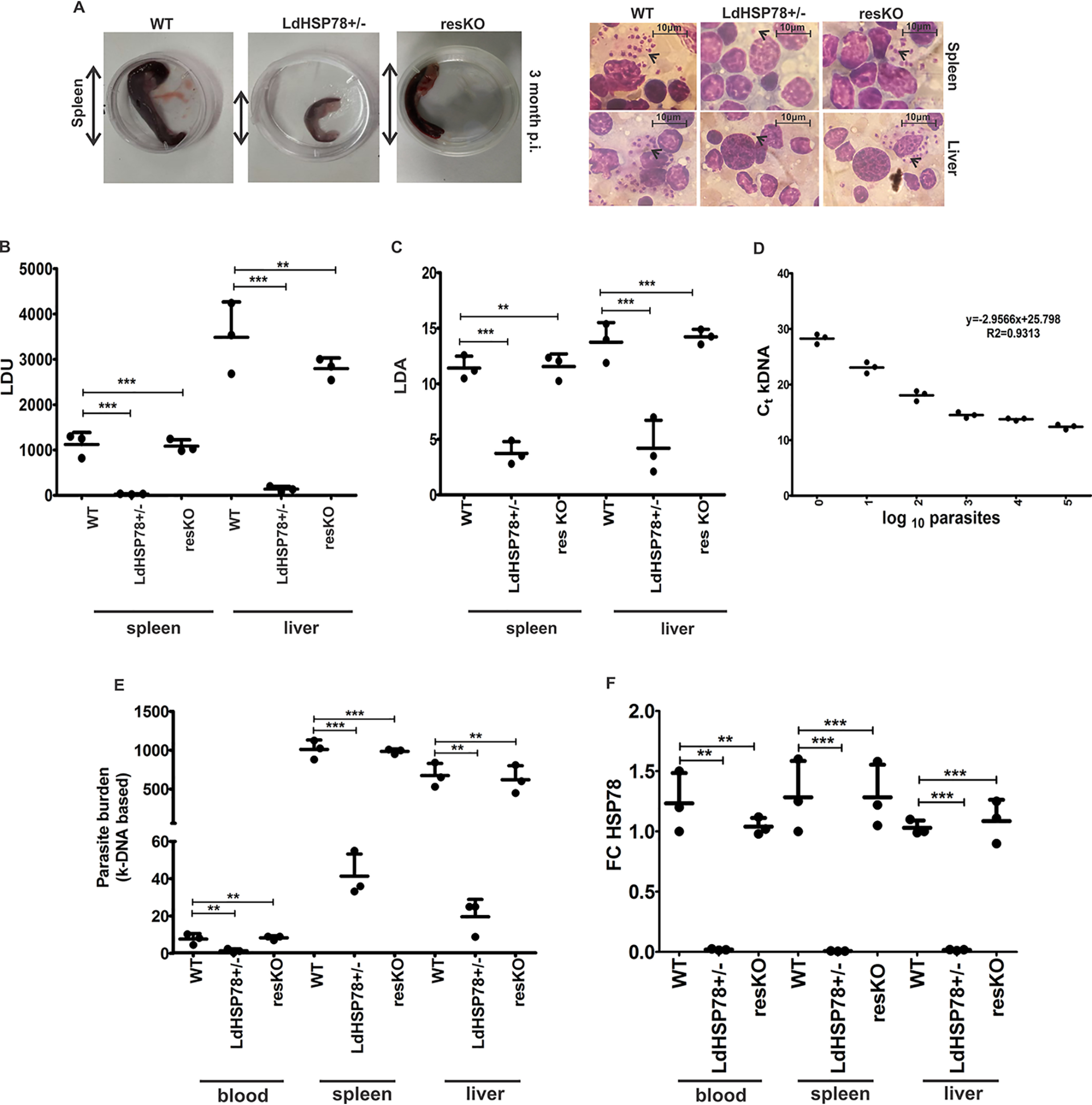
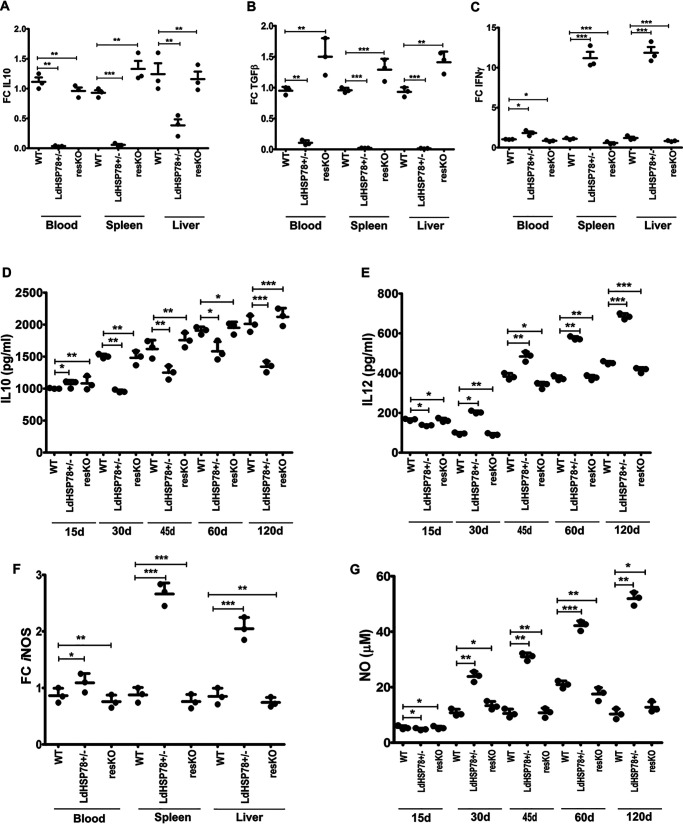
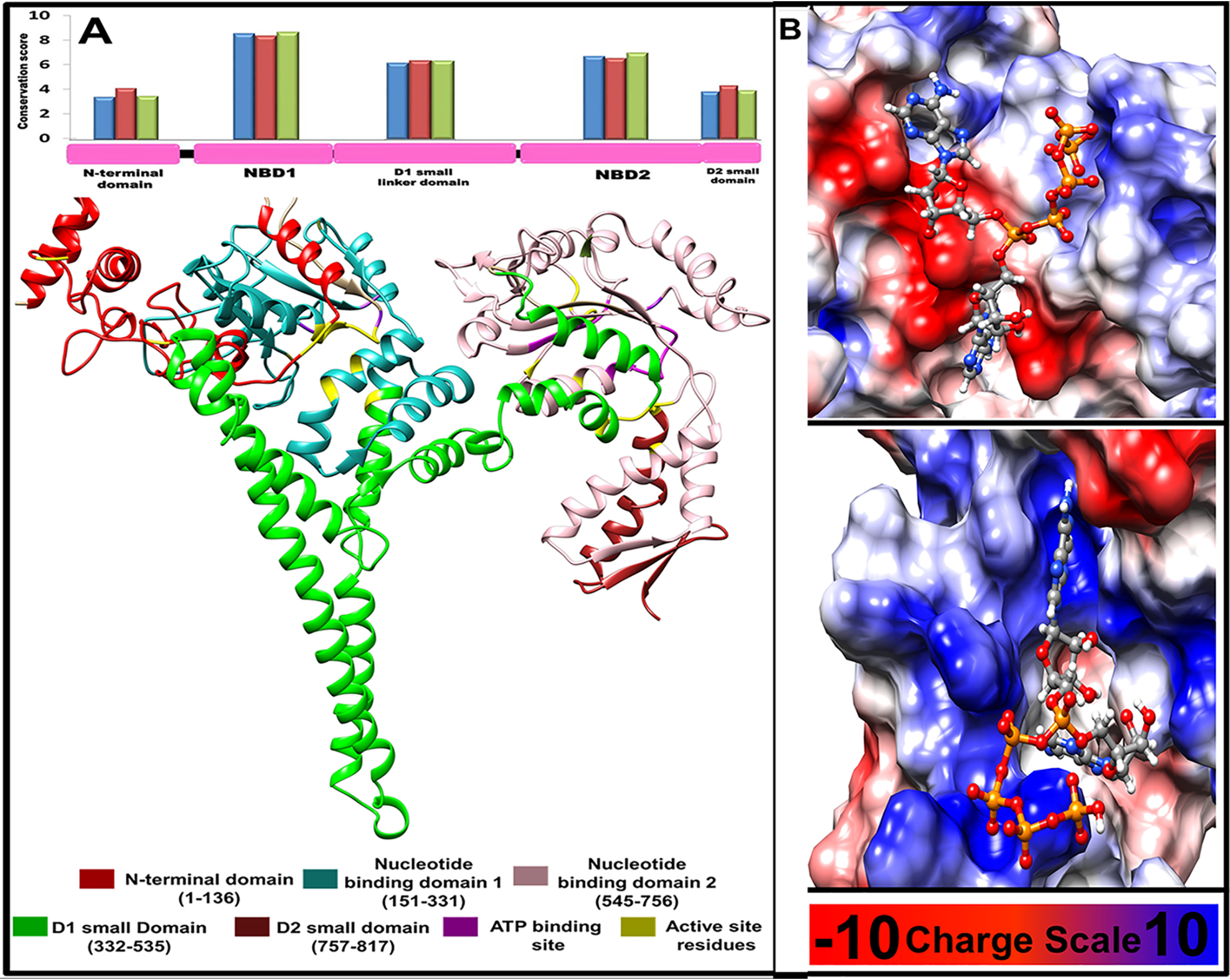
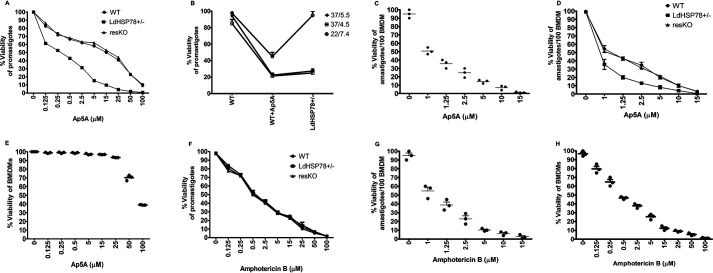
The emergence of resistance to available antileishmanial drugs advocates identification of new drug targets and their inhibitors for visceral leishmaniasis. Here, we identified Leishmania donovani heat shock protein 78 (LdHSP78), a putative caseinolytic protease, as important for parasite infection of host macrophages and a potential therapeutic target. Enrichment of LdHSP78 in infected humans, hamsters, and parasite amastigotes suggested its importance for disease persistence. Heterozygous knockouts of L. donovani HSP78 (LdHSP78+/-) and Leishmania mexicana HSP78 (LmxHSP78+/-) were generated using a flanking UTR-based multifragment ligation strategy and the CRISPR-Cas9 technique, respectively to investigate the significance of HSP78 for disease manifestation. The LdHSP78+/- parasite burden was dramatically reduced in both murine bone marrow-derived macrophages and hamsters, in association with enrichment of proinflammatory cytokines and NO. This finding implies that LdHSP78+/- parasites cannot suppress immune activation and escape NO-mediated toxicity in macrophages. Furthermore, phosphorylation of the mitogen-activated protein kinase p38 was enhanced and phosphorylation of extracellular signal-regulated kinase 1/2 was decreased in cells infected with LdHSP78+/- parasites, compared with WT parasites. Virulence of the LdHSP78+/- strain was restored by episomal addition of the LdHSP78 gene. Finally, using high-throughput virtual screening, we identified P1,P5-di(adenosine-5')-pentaphosphate (Ap5A) ammonium salt as an LdHSP78 inhibitor. It selectively induced amastigote death at doses similar to amphotericin B doses, while exhibiting much less cytotoxicity to healthy macrophages than amphotericin B. In summary, using both a genetic knockout approach and pharmacological inhibition, we establish LdHSP78 as a drug target and Ap5A as a potential lead for improved antileishmanial agents.
Keywords: Ap5A; Leishmania; caseinolytic protease; drug screening; heat shock protein 78 (HSP78); high-throughput virtual screening; innate immunity; kinase signaling; macrophage; molecular modeling; transgenic parasite.
© 2020 Das et al.
Conflict of interest statement
Conflict of interest—The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Pallavi R., Roy N., Nageshan R. K., Talukdar P., Pavithra S. R., Reddy R., Venketesh S., Kumar R., Gupta A. K., Singh R. K., Yadav S. C., and Tatu U. (2010) Heat shock protein 90 as a drug target against protozoan infections: biochemical characterization of HSP90 from Plasmodium falciparum and Trypanosoma evansi and evaluation of its inhibitor as a candidate drug. J. Biol. Chem. 285, 37964–37975 10.1074/jbc.M110.155317 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
