

Pre-existing antibody-mediated adverse effects prevent the clinical development of a bacterial anti-inflammatory protein
- PMID: 32471891
- PMCID: PMC7541340
- DOI: 10.1242/dmm.045534
Pre-existing antibody-mediated adverse effects prevent the clinical development of a bacterial anti-inflammatory protein
Abstract
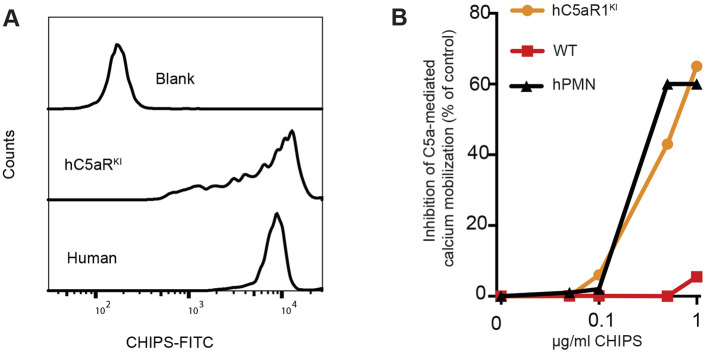
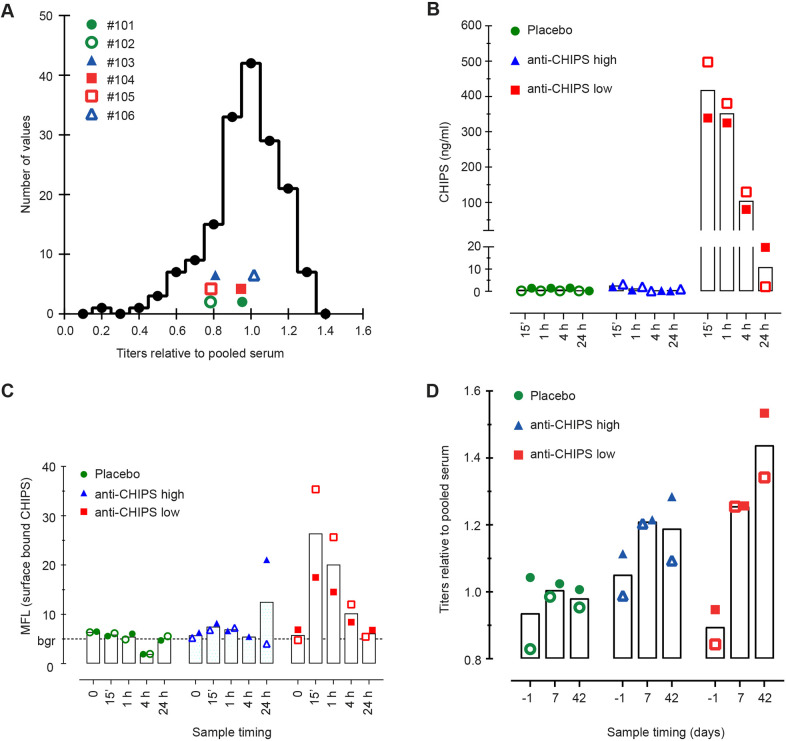
Bacterial pathogens have evolved to secrete strong anti-inflammatory proteins that target the immune system. It was long speculated whether these virulence factors could serve as therapeutics in diseases in which abnormal immune activation plays a role. We adopted the secreted chemotaxis inhibitory protein of Staphylococcus aureus (CHIPS) as a model virulence factor-based therapeutic agent for diseases in which C5AR1 stimulation plays an important role. We show that the administration of CHIPS in human C5AR1 knock-in mice successfully dampens C5a-mediated neutrophil migration during immune complex-initiated inflammation. Subsequent CHIPS toxicology studies in animal models were promising. However, during a small phase I trial, healthy human volunteers showed adverse effects directly after CHIPS administration. Subjects showed clinical signs of anaphylaxis with mild leukocytopenia and increased C-reactive protein concentrations, which are possibly related to the presence of relatively high circulating anti-CHIPS antibodies and suggest an inflammatory response. Even though our data in mice show CHIPS as a potential anti-inflammatory agent, safety issues in human subjects temper the use of CHIPS in its current form as a therapeutic candidate. The use of staphylococcal proteins, or other bacterial proteins, as therapeutics or immune-modulators in humans is severely hampered by pre-existing circulating antibodies.
Keywords: C5aR chemotaxis; CHIPS; Clinical trials; Humanized mouse; Immune complex.
© 2020. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures



References
-
- Bekker P., Dairaghi D., Seitz L., Leleti M., Wang Y., Ertl L., Baumgart T., Shugarts S., Lohr L., Dang T. et al. (2016). Characterization of pharmacologic and pharmacokinetic properties of CCX168, a potent and selective orally administered complement 5a receptor inhibitor, based on preclinical evaluation and randomized phase 1 clinical study. PLoS ONE 11, e0164646 10.1371/journal.pone.0164646 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
