

Crosstalk between androgen and Wnt/β-catenin leads to changes of wool density in FGF5-knockout sheep
- PMID: 32472005
- PMCID: PMC7260202
- DOI: 10.1038/s41419-020-2622-x
Crosstalk between androgen and Wnt/β-catenin leads to changes of wool density in FGF5-knockout sheep
Abstract
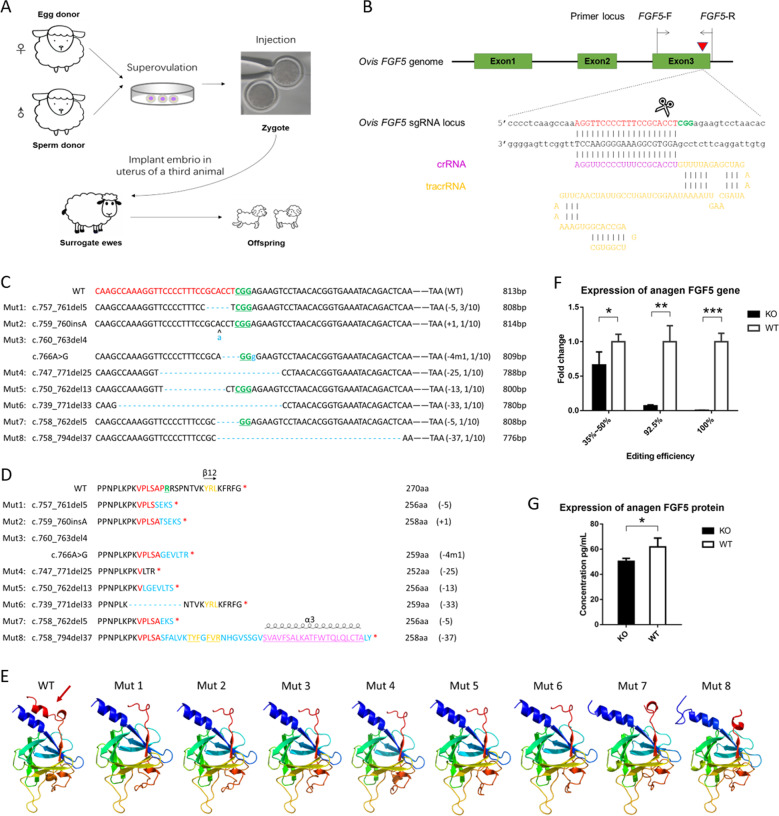
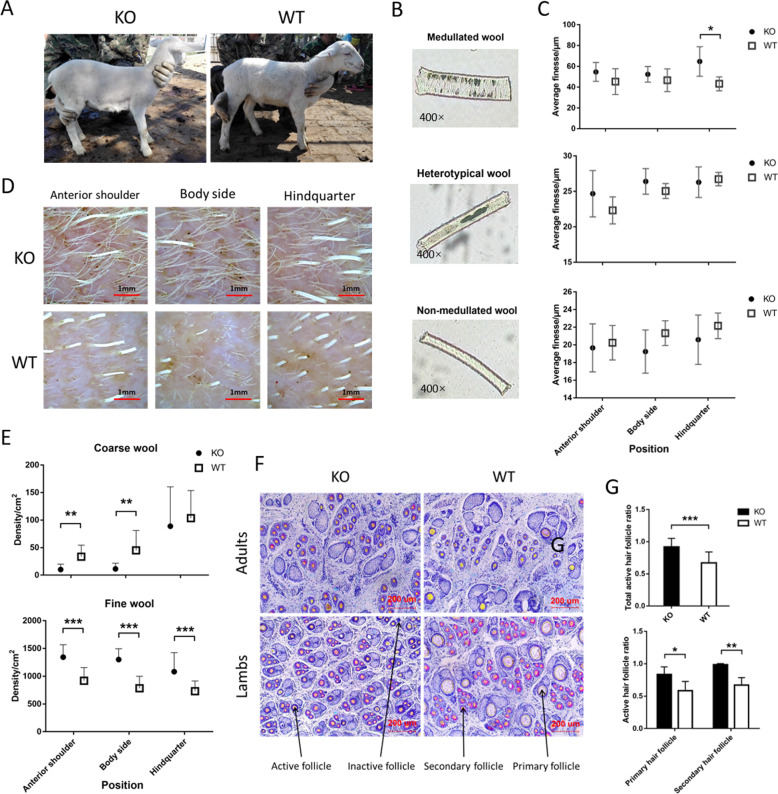
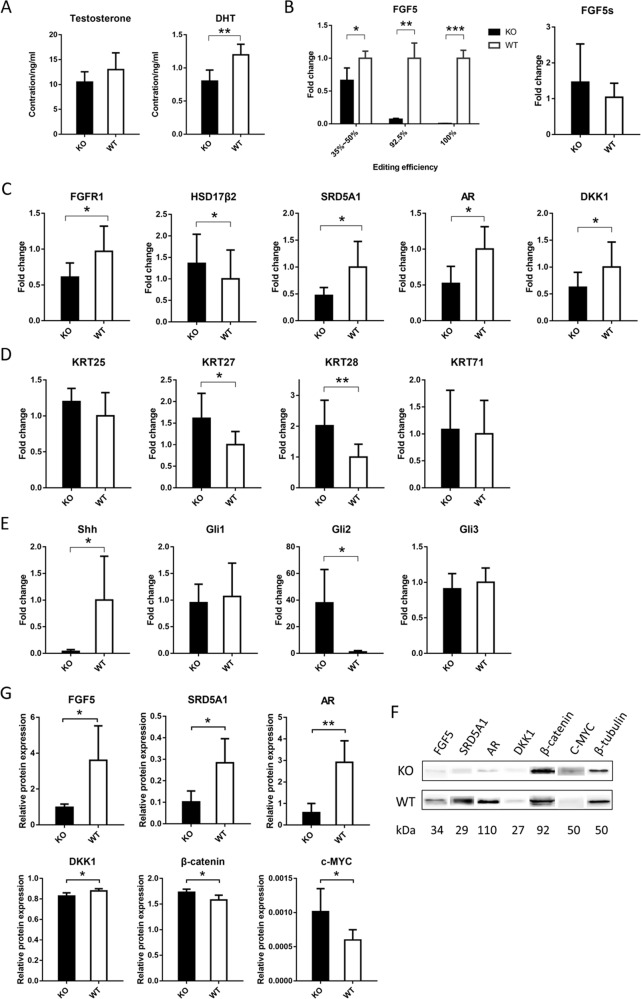
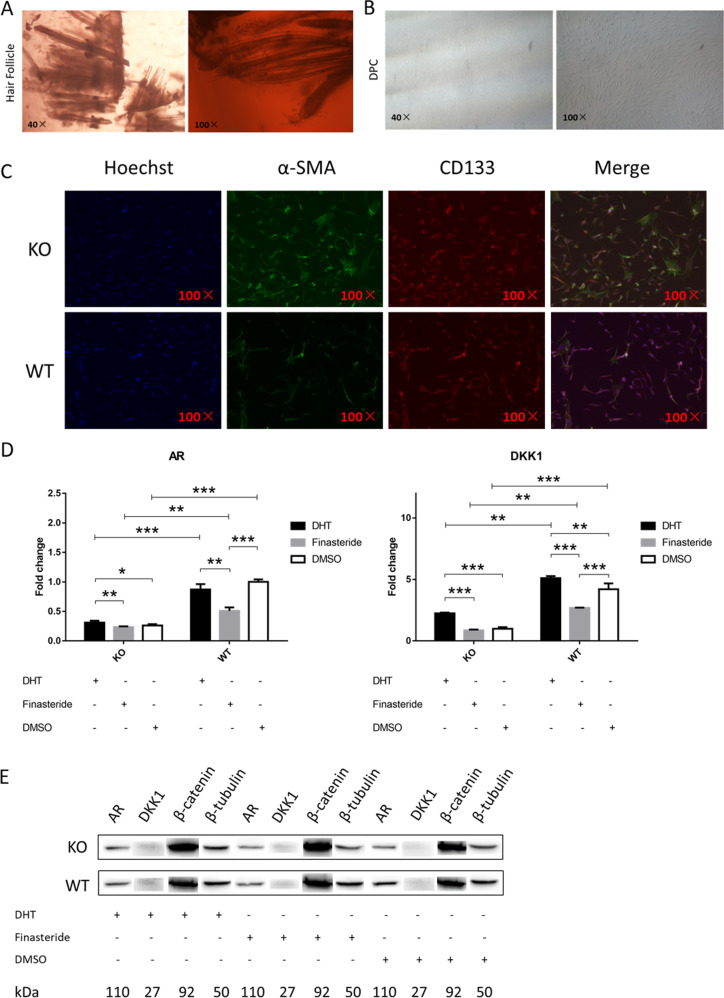
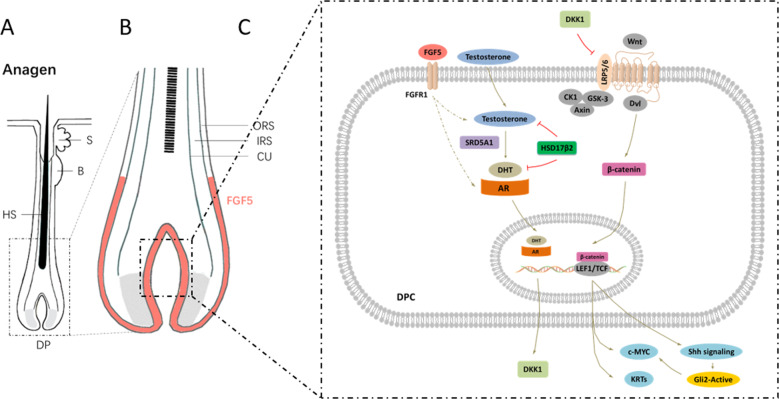
Fibroblast growth factor 5 (FGF5) is a famous dominant inhibitor of anagen phase of hair cycle. Mutations of FGF5 gene result in a longer wool in mice, donkeys, dogs, cats, and even in human eyelashes. Sheep is an important source of wool production. How to improve the production of wool quickly and effectively is an urgent problem to be solved. In this study, we generated five FGF5-knockout Dorper sheep by the CRISPR/Cas9 system. The expression level of FGF5 mRNA in knockout (KO) sheep decreased significantly, and all FGF5 proteins were dysfunctional. The KO sheep displayed a significant increase in fine-wool and active hair-follicle density. The crosstalk between androgen and Wnt/β-catenin signaling downstream of FGF5 gene plays a key role. We established downstream signaling cascades for the first time, including FGF5, FGFR1, androgen, AR, Wnt/β-catenin, Shh/Gli2, c-MYC, and KRTs. These findings further improved the function of FGF5 gene, and provided therapeutic ideas for androgen alopecia.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Increasing GSH-Px Activity and Activating Wnt Pathway Promote Fine Wool Growth in FGF5-Edited Sheep.Cells. 2024 Jun 5;13(11):985. doi: 10.3390/cells13110985. Cells. 2024. PMID: 38891117 Free PMC article.
-
RAPID COMMUNICATION: Generation of FGF5 knockout sheep via the CRISPR/Cas9 system.J Anim Sci. 2017 May;95(5):2019-2024. doi: 10.2527/jas.2017.1503. J Anim Sci. 2017. PMID: 28727005
-
Bioinformatics analysis of evolutionary characteristics and biochemical structure of FGF5 Gene in sheep.Gene. 2019 Jun 20;702:123-132. doi: 10.1016/j.gene.2019.03.040. Epub 2019 Mar 26. Gene. 2019. PMID: 30926307
-
Androgen modulation of Wnt/β-catenin signaling in androgenetic alopecia.Arch Dermatol Res. 2018 Jul;310(5):391-399. doi: 10.1007/s00403-018-1826-8. Epub 2018 Mar 16. Arch Dermatol Res. 2018. PMID: 29549490 Review.
-
Androgen actions on the human hair follicle: perspectives.Exp Dermatol. 2013 Mar;22(3):168-71. doi: 10.1111/exd.12024. Epub 2012 Sep 28. Exp Dermatol. 2013. PMID: 23016593 Review.
Cited by
-
Gender-Difference in Hair Length as Revealed by Crispr-Based Production of Long-Haired Mice with Dysfunctional FGF5 Mutations.Int J Mol Sci. 2022 Oct 6;23(19):11855. doi: 10.3390/ijms231911855. Int J Mol Sci. 2022. PMID: 36233155 Free PMC article.
-
Genetics of the phenotypic evolution in sheep: a molecular look at diversity-driving genes.Genet Sel Evol. 2022 Sep 9;54(1):61. doi: 10.1186/s12711-022-00753-3. Genet Sel Evol. 2022. PMID: 36085023 Free PMC article. Review.
-
Construction of a high-density genetic map and QTL localization of body weight and wool production related traits in Alpine Merino sheep based on WGR.BMC Genomics. 2024 Jun 27;25(1):641. doi: 10.1186/s12864-024-10535-4. BMC Genomics. 2024. PMID: 38937677 Free PMC article.
-
Recent Advances in the Role of Fibroblast Growth Factors in Hair Follicle Growth.Biomolecules. 2025 Aug 20;15(8):1198. doi: 10.3390/biom15081198. Biomolecules. 2025. PMID: 40867641 Free PMC article. Review.
-
Opportunities for CRISPR-Cas9 application in farm animal genetic improvement.Mol Biol Rep. 2024 Oct 30;51(1):1108. doi: 10.1007/s11033-024-10052-3. Mol Biol Rep. 2024. PMID: 39476174 Review.
References
-
- Stenn KS, Paus R. Controls of hair follicle cycling. Physiol. Rev. 2001;81:449. - PubMed
-
- Schneider MR, Schmidt-Ullrich R, Paus R. The hair follicle as a dynamic miniorgan. Curr. Biol. 2009;19:R132. - PubMed
-
- Krause K, Foitzik K. Biology of the hair follicle: the basics. Semin. Cutan. Med. Surg. 2006;25:2. - PubMed
-
- Fuchs, E., Merrill, B. J., Jamora, C. & DasGupta, R. At the Roots of a Never-Ending Cycle. Dev. Cell1, 13–25 (2001). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
