

Studying RNA-DNA interactome by Red-C identifies noncoding RNAs associated with various chromatin types and reveals transcription dynamics
- PMID: 32479626
- PMCID: PMC7337940
- DOI: 10.1093/nar/gkaa457
Studying RNA-DNA interactome by Red-C identifies noncoding RNAs associated with various chromatin types and reveals transcription dynamics
Abstract
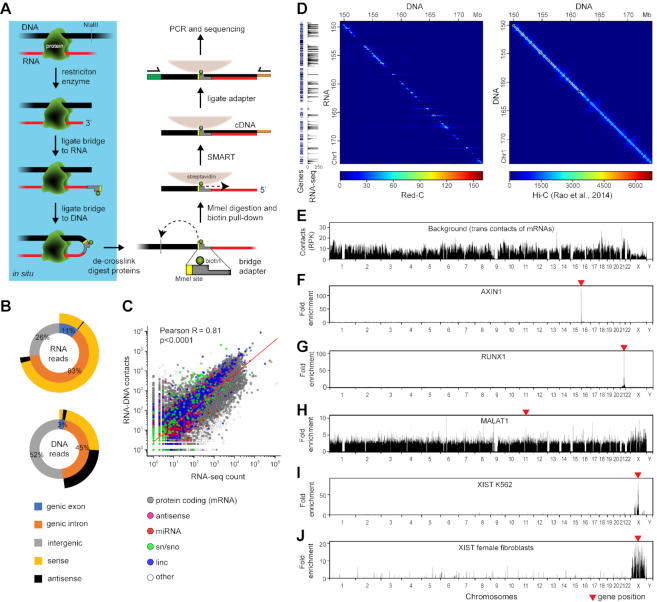
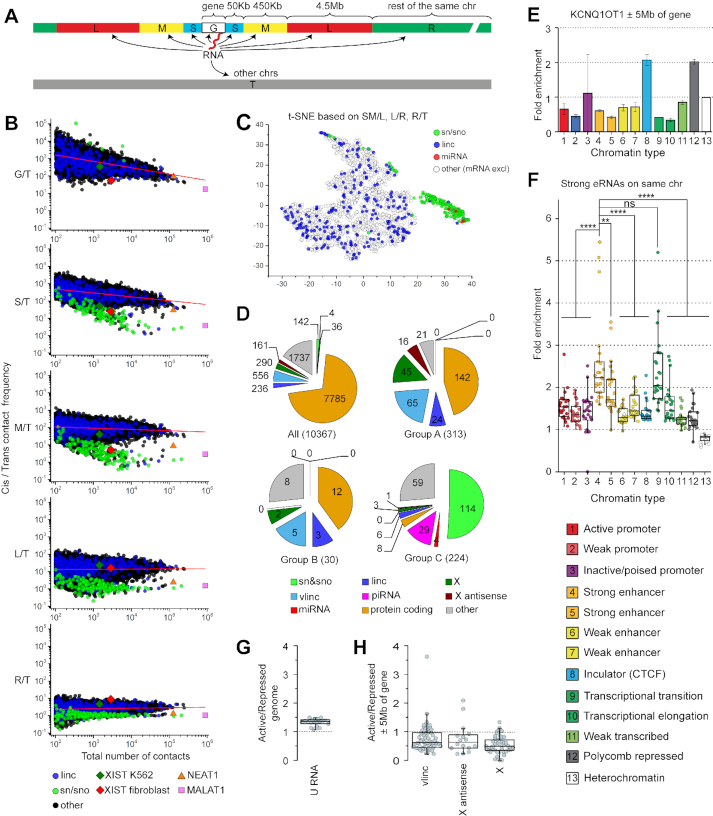
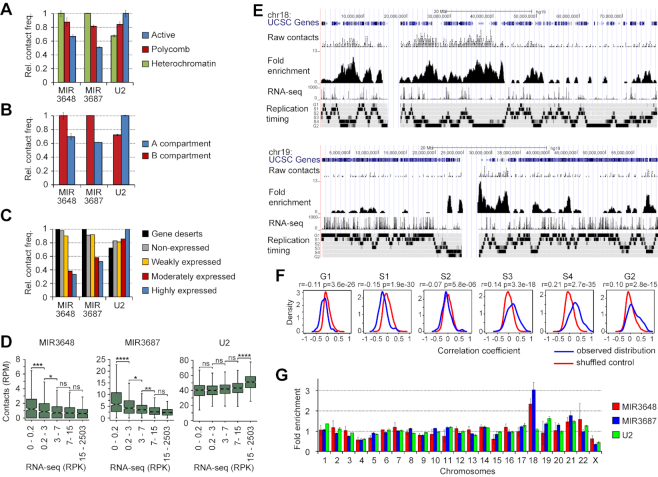
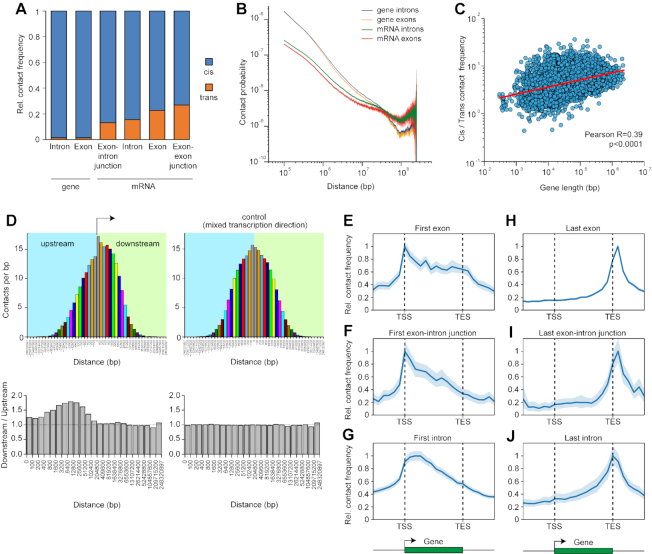
Non-coding RNAs (ncRNAs) participate in various biological processes, including regulating transcription and sustaining genome 3D organization. Here, we present a method termed Red-C that exploits proximity ligation to identify contacts with the genome for all RNA molecules present in the nucleus. Using Red-C, we uncovered the RNA-DNA interactome of human K562 cells and identified hundreds of ncRNAs enriched in active or repressed chromatin, including previously undescribed RNAs. Analysis of the RNA-DNA interactome also allowed us to trace the kinetics of messenger RNA production. Our data support the model of co-transcriptional intron splicing, but not the hypothesis of the circularization of actively transcribed genes.
© The Author(s) 2020. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Kanehisa T., Tanaka T., Kano Y.. Low molecular RNA associated with chromatin: purification and characterization of RNA that stimulates RNA synthesis. Biochim. Biophys. Acta. 1972; 277:584–589. - PubMed
-
- Holmes D.S., Mayfield J.E., Sander G., Bonner J.. Chromosomal RNA: its properties. Science. 1972; 177:72–74. - PubMed
-
- Bynum J.W., Volkin E.. Chromatin-associated RNA: differential extraction and characterization. Biochim. Biophys. Acta. 1980; 607:304–318. - PubMed
-
- Nozawa R.S., Gilbert N.. RNA: nuclear glue for folding the genome. Trends Cell Biol. 2019; 29:201–211. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
