

Liquid network connectivity regulates the stability and composition of biomolecular condensates with many components
- PMID: 32482873
- PMCID: PMC7306995
- DOI: 10.1073/pnas.1917569117
Liquid network connectivity regulates the stability and composition of biomolecular condensates with many components
Abstract
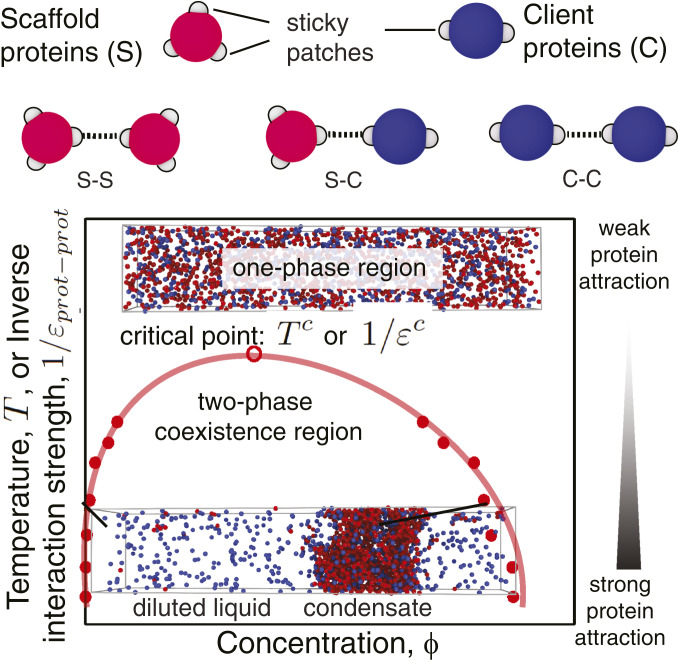
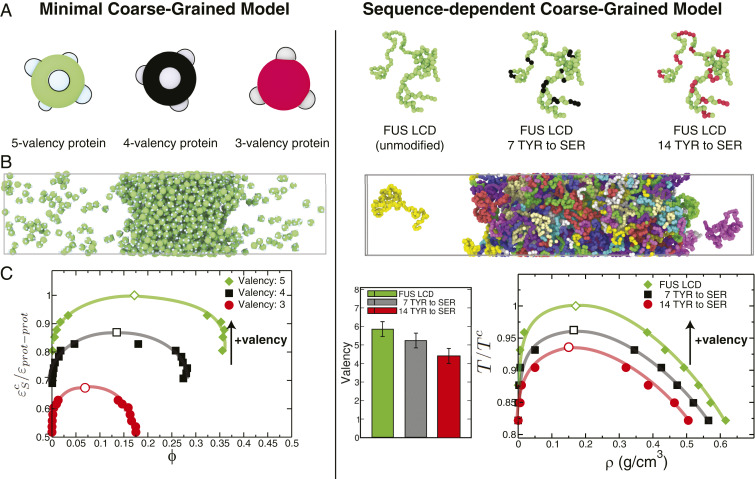
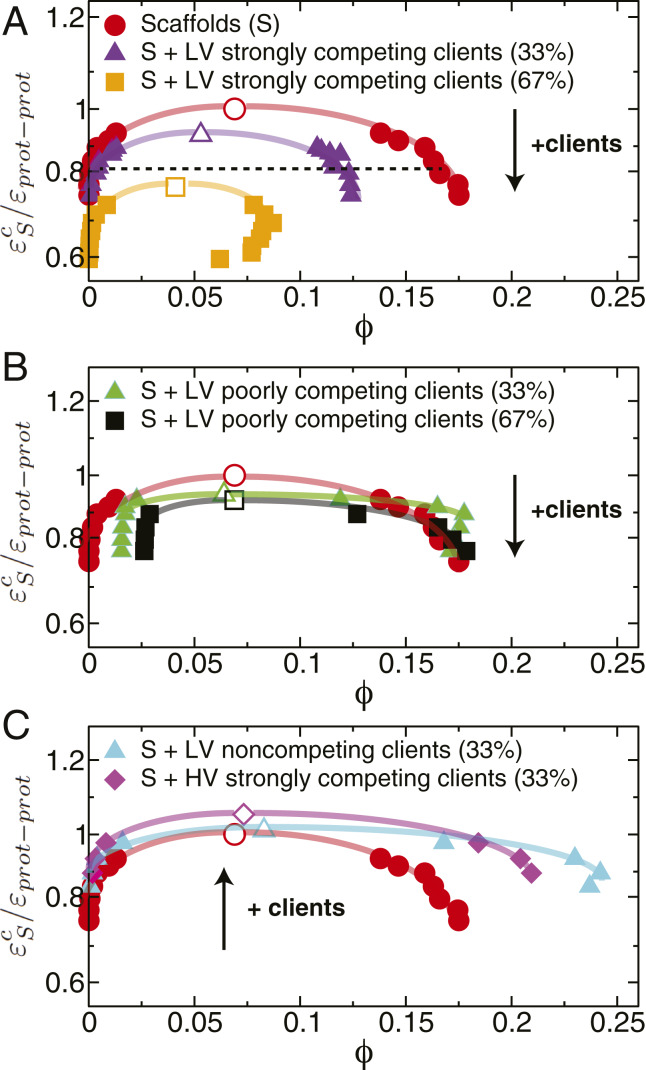
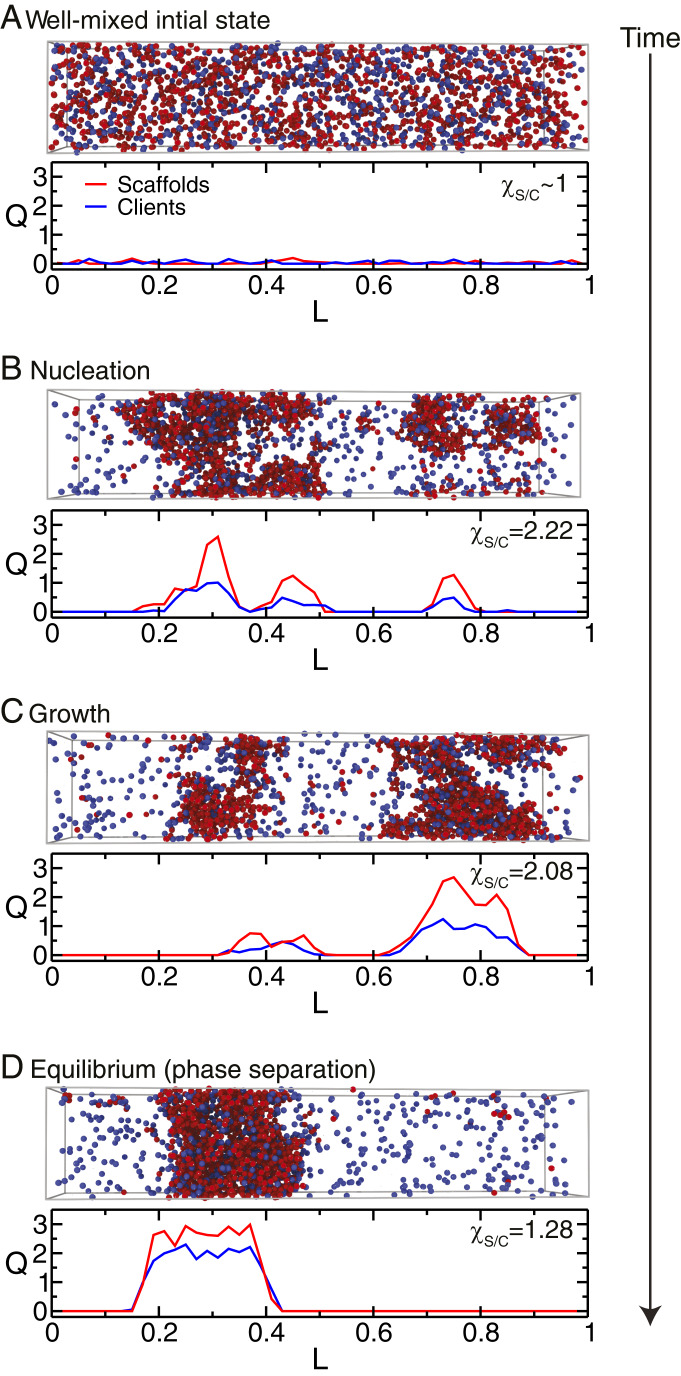
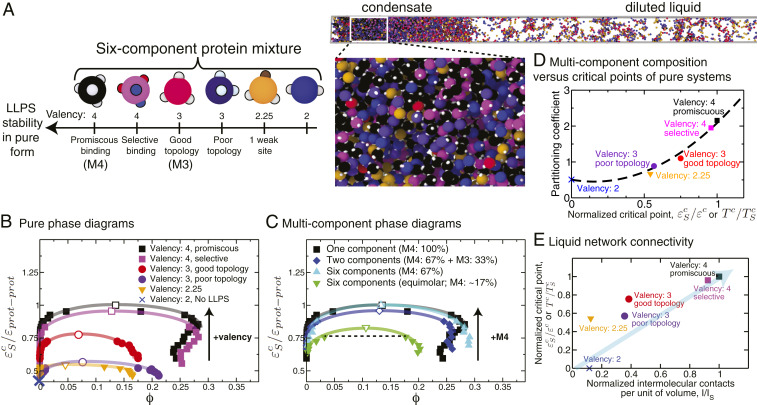
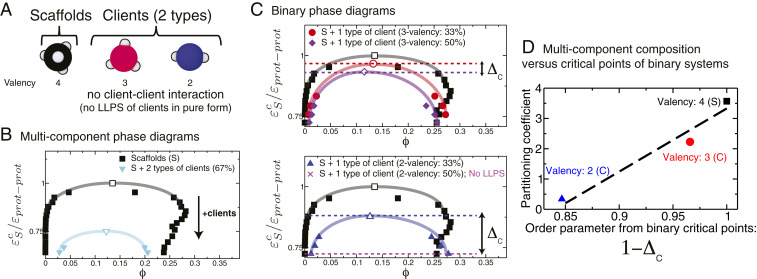
One of the key mechanisms used by cells to control the spatiotemporal organization of their many components is the formation and dissolution of biomolecular condensates through liquid-liquid phase separation (LLPS). Using a minimal coarse-grained model that allows us to simulate thousands of interacting multivalent proteins, we investigate the physical parameters dictating the stability and composition of multicomponent biomolecular condensates. We demonstrate that the molecular connectivity of the condensed-liquid network-i.e., the number of weak attractive protein-protein interactions per unit of volume-determines the stability (e.g., in temperature, pH, salt concentration) of multicomponent condensates, where stability is positively correlated with connectivity. While the connectivity of scaffolds (biomolecules essential for LLPS) dominates the phase landscape, introduction of clients (species recruited via scaffold-client interactions) fine-tunes it by transforming the scaffold-scaffold bond network. Whereas low-valency clients that compete for scaffold-scaffold binding sites decrease connectivity and stability, those that bind to alternate scaffold sites not required for LLPS or that have higher-than-scaffold valencies form additional scaffold-client-scaffold bridges increasing stability. Proteins that establish more connections (via increased valencies, promiscuous binding, and topologies that enable multivalent interactions) support the stability of and are enriched within multicomponent condensates. Importantly, proteins that increase the connectivity of multicomponent condensates have higher critical points as pure systems or, if pure LLPS is unfeasible, as binary scaffold-client mixtures. Hence, critical points of accessible systems (i.e., with just a few components) might serve as a unified thermodynamic parameter to predict the composition of multicomponent condensates.
Keywords: biomolecular condensates; cell compartmentalization; liquid–liquid phase separation; membraneless organelles.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Sear R. P., The cytoplasm of living cells: A functional mixture of thousands of components. J. Phys. Condens. Matter 17, S3587–S3595 (2005).
-
- Alberts B., Molecular Biology of the Cell (Garland Science, Taylor and Francis Group, New York, NY, ed. 6, 2015).
-
- Hyman A. A., Weber C. A., Jülicher F., Liquid-liquid phase separation in biology. Annu. Rev. Cell Dev. Biol. 30, 39–58 (2014). - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
