

The polycomb group protein PCGF6 mediates germline gene silencing by recruiting histone-modifying proteins to target gene promoters
- PMID: 32482889
- PMCID: PMC7363147
- DOI: 10.1074/jbc.RA119.012121
The polycomb group protein PCGF6 mediates germline gene silencing by recruiting histone-modifying proteins to target gene promoters
Abstract
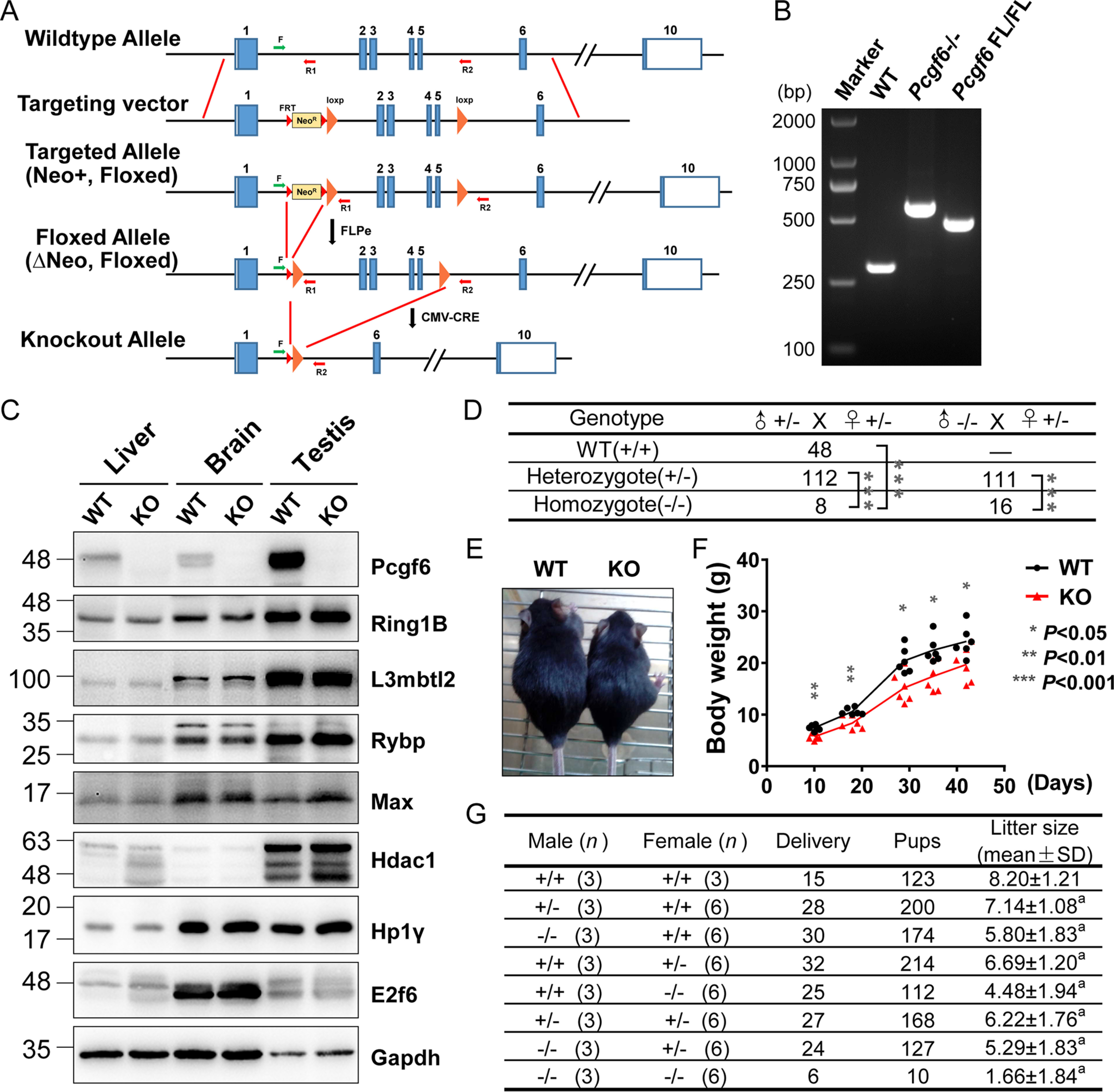
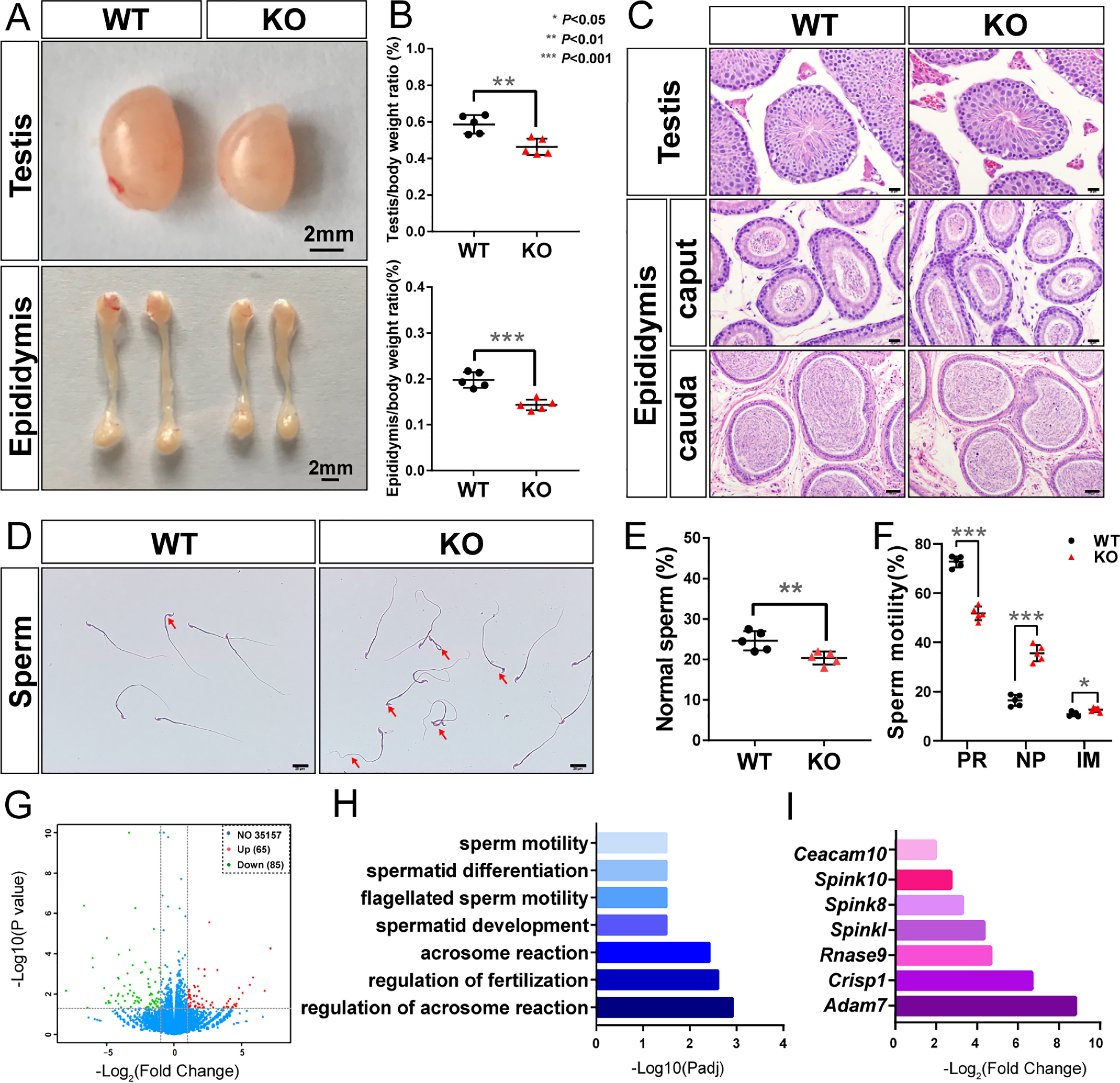
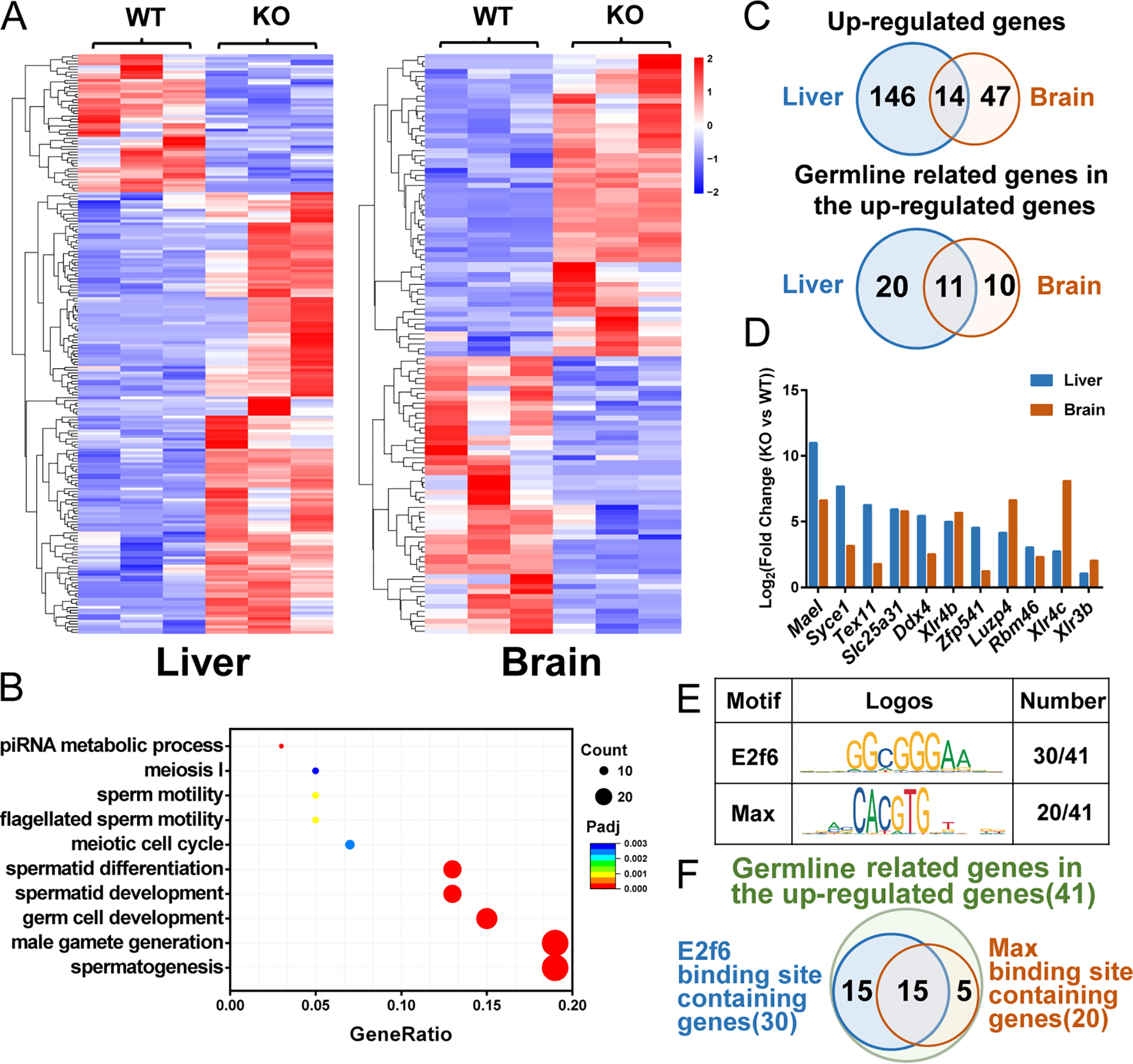
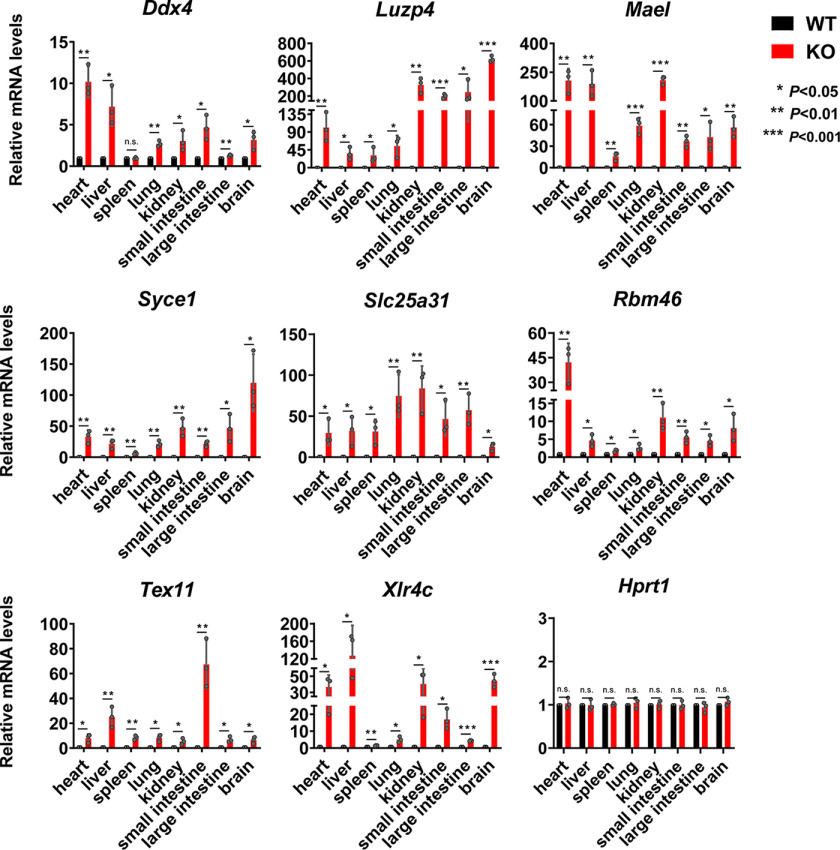
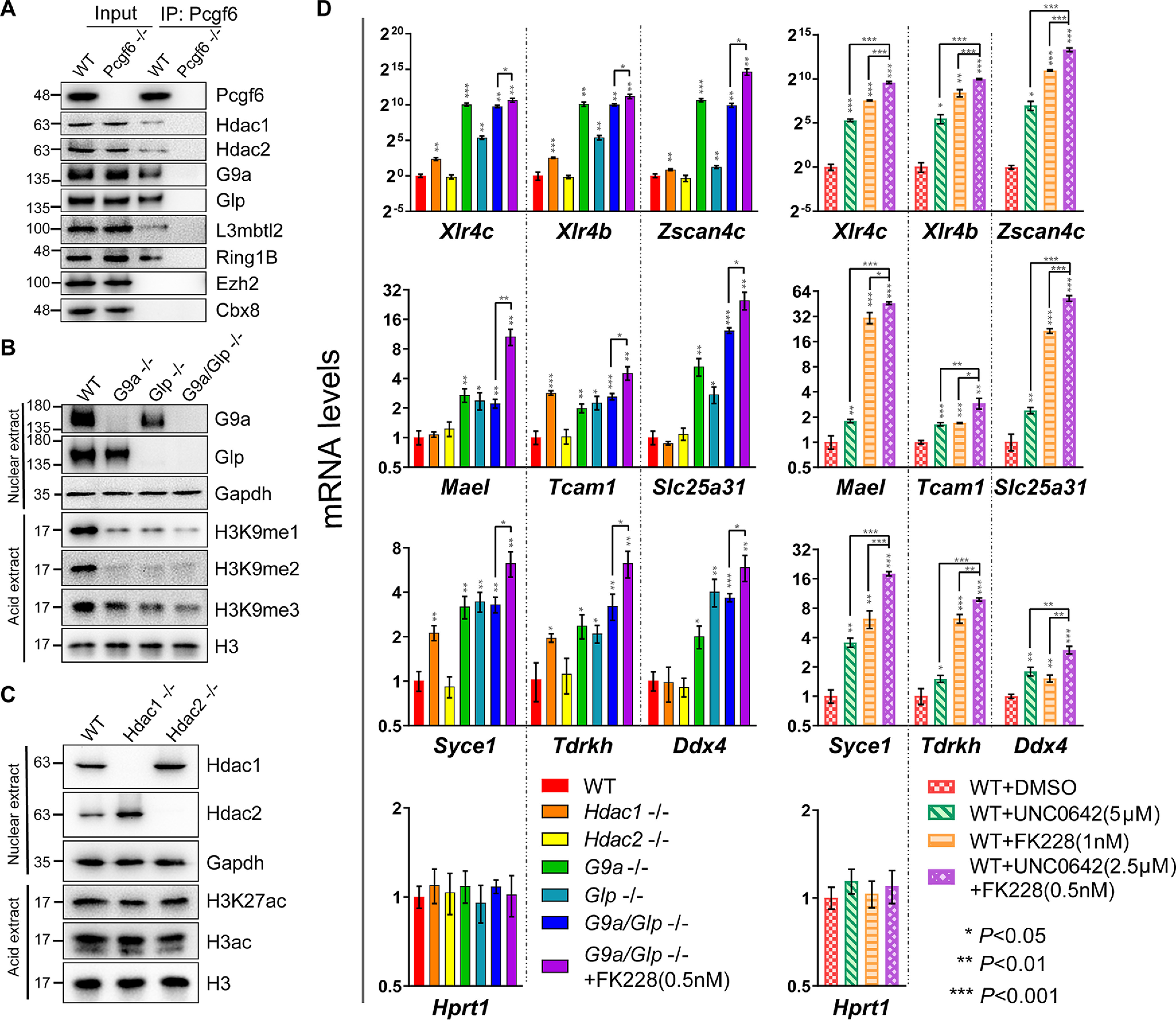
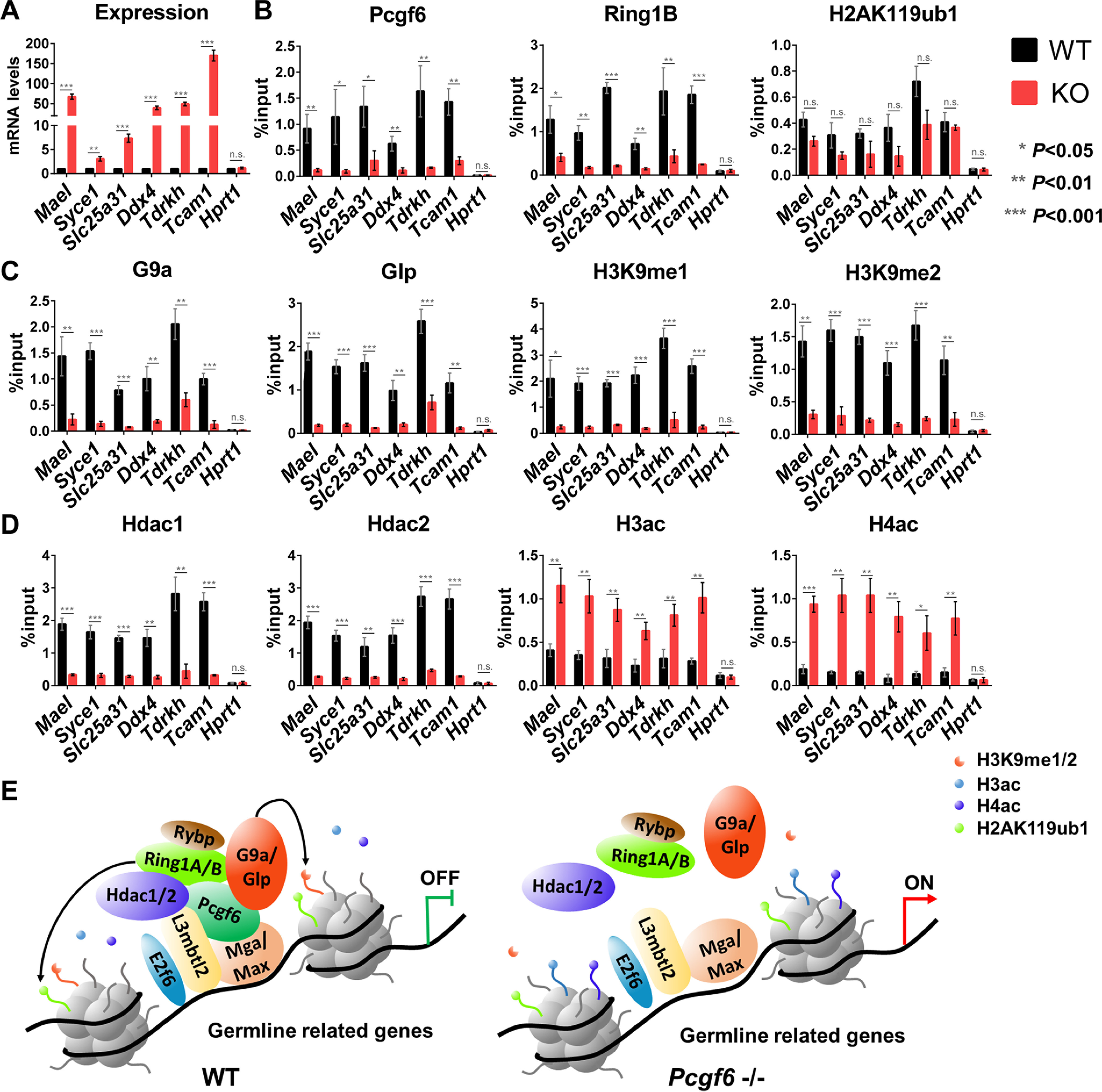
Polycomb group (PcG) proteins are essential for maintenance of lineage fidelity by coordinating developmental gene expression programs. Polycomb group ring finger 6 (PCGF6) has been previously reported to repress expression of lineage-specific genes, especially germ cell-related genes in mouse embryonic stem cells (ESCs) via the noncanonical polycomb repressive complex PRC1.6. However, the molecular mechanism of this repression remains largely unknown. Here, using RNA-Seq, real-time RT-PCR, immunohistochemistry, immunoprecipitation, and ChIP analyses, we demonstrate that PCGF6 plays an essential role in embryonic development, indicated by the partially penetrant embryonic lethality in homozygous PCGF6 (Pcgf6-/-)-deficient mice. We also found that surviving Pcgf6-deficient mice exhibit reduced fertility. Using the Pcgf6-deficient mice, we observed that ablation of Pcgf6 in somatic tissues robustly derepresses germ cell-related genes. We further provide evidence that these genes are direct targets of PCGF6 in ESCs and that endogenous PCGF6 co-localizes with the histone-modifying proteins G9A histone methyltransferase (G9A)/G9a-like protein (GLP) and histone deacetylase 1/2 (HDAC1/2) on the promoters of the germ cell-related genes. Moreover, the binding of these proteins to their target genes correlated with methylation of Lys-9 of histone 3 and with the status of histone acetylation at these genes. Moreover, the recruitment of G9A/GLP and HDAC1/2 to target promoters depended on the binding of PCGF6. Our findings indicate that PCGF6 has a critical role in safeguarding lineage decisions and in preventing aberrant expression of germ cell-related genes.
Keywords: G9A histone methyltransferase; G9a-like protein 1 (Glp); G9a/Glp; PRC1.6; Pcgf6; embryonic stem cell; embryonic stem cells; epigenetic regulation; gene knockout; gene regulation; germline gene silencing; histone deacetylase (HDAC); histone deacetylase 1/2 (Hdac1/2); histone methylation; knockout mice; polycomb; polycomb group ring finger 6 (Pcgf6).
© 2020 Liu et al.
Conflict of interest statement
Conflict of interest—The authors declare that they have no conflicts of interest with the contents of this article.
Figures
Similar articles
-
PCGF6-PRC1 suppresses premature differentiation of mouse embryonic stem cells by regulating germ cell-related genes.Elife. 2017 Mar 17;6:e21064. doi: 10.7554/eLife.21064. Elife. 2017. PMID: 28304275 Free PMC article.
-
DNMTs and SETDB1 function as co-repressors in MAX-mediated repression of germ cell-related genes in mouse embryonic stem cells.PLoS One. 2018 Nov 7;13(11):e0205969. doi: 10.1371/journal.pone.0205969. eCollection 2018. PLoS One. 2018. PMID: 30403691 Free PMC article.
-
Pcgf6, a polycomb group protein, regulates mesodermal lineage differentiation in murine ESCs and functions in iPS reprogramming.Stem Cells. 2014 Dec;32(12):3112-25. doi: 10.1002/stem.1826. Stem Cells. 2014. PMID: 25187489
-
Polycomb complexes in stem cells and embryonic development.Development. 2013 Jun;140(12):2525-34. doi: 10.1242/dev.091553. Development. 2013. PMID: 23715546 Review.
-
Pcgf5 Contributes to PRC1 (Polycomb Repressive Complex 1) in Developing Cardiac Cells.2016 Jun 25. In: Nakanishi T, Markwald RR, Baldwin HS, Keller BB, Srivastava D, Yamagishi H, editors. Etiology and Morphogenesis of Congenital Heart Disease: From Gene Function and Cellular Interaction to Morphology [Internet]. Tokyo: Springer; 2016. Chapter 43. 2016 Jun 25. In: Nakanishi T, Markwald RR, Baldwin HS, Keller BB, Srivastava D, Yamagishi H, editors. Etiology and Morphogenesis of Congenital Heart Disease: From Gene Function and Cellular Interaction to Morphology [Internet]. Tokyo: Springer; 2016. Chapter 43. PMID: 29787140 Free Books & Documents. Review.
Cited by
-
Identification of candidate genes associated with bacterial and viral infections in wild boars hunted in Tuscany (Italy).Sci Rep. 2022 May 17;12(1):8145. doi: 10.1038/s41598-022-12353-8. Sci Rep. 2022. PMID: 35581286 Free PMC article.
-
An Epigenomic Roadmap Primes Non-Growing Oocytes for Maturation and Early Embryogenesis.bioRxiv [Preprint]. 2025 Jul 6:2025.07.06.663377. doi: 10.1101/2025.07.06.663377. bioRxiv. 2025. PMID: 40631288 Free PMC article. Preprint.
-
Repression of germline genes by PRC1.6 and SETDB1 in the early embryo precedes DNA methylation-mediated silencing.Nat Commun. 2021 Dec 2;12(1):7020. doi: 10.1038/s41467-021-27345-x. Nat Commun. 2021. PMID: 34857746 Free PMC article.
-
E2F6 initiates stable epigenetic silencing of germline genes during embryonic development.Nat Commun. 2021 Jun 11;12(1):3582. doi: 10.1038/s41467-021-23596-w. Nat Commun. 2021. PMID: 34117224 Free PMC article.
-
A rapid and stable spontaneous reprogramming system of Spermatogonial stem cells to Pluripotent State.Cell Biosci. 2023 Dec 1;13(1):222. doi: 10.1186/s13578-023-01150-z. Cell Biosci. 2023. PMID: 38041111 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
