

Integrative omics analysis of the termite gut system adaptation to Miscanthus diet identifies lignocellulose degradation enzymes
- PMID: 32483294
- PMCID: PMC7264248
- DOI: 10.1038/s42003-020-1004-3
Integrative omics analysis of the termite gut system adaptation to Miscanthus diet identifies lignocellulose degradation enzymes
Abstract
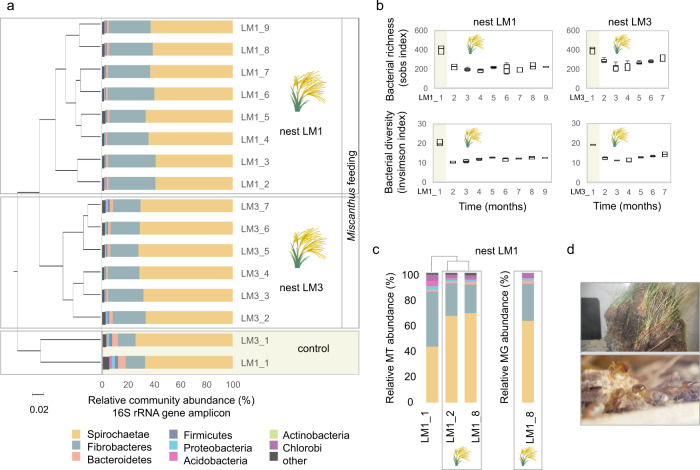
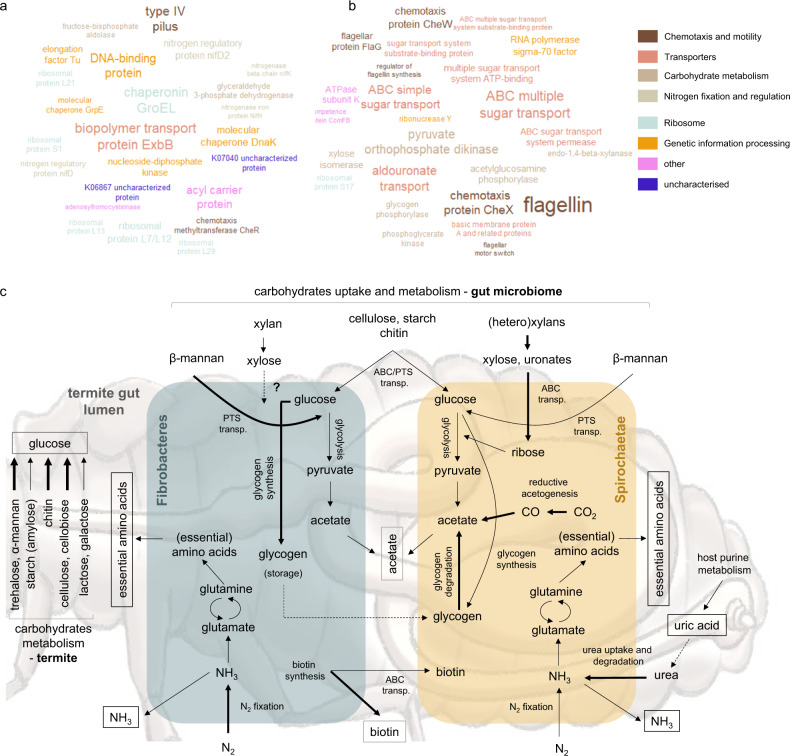
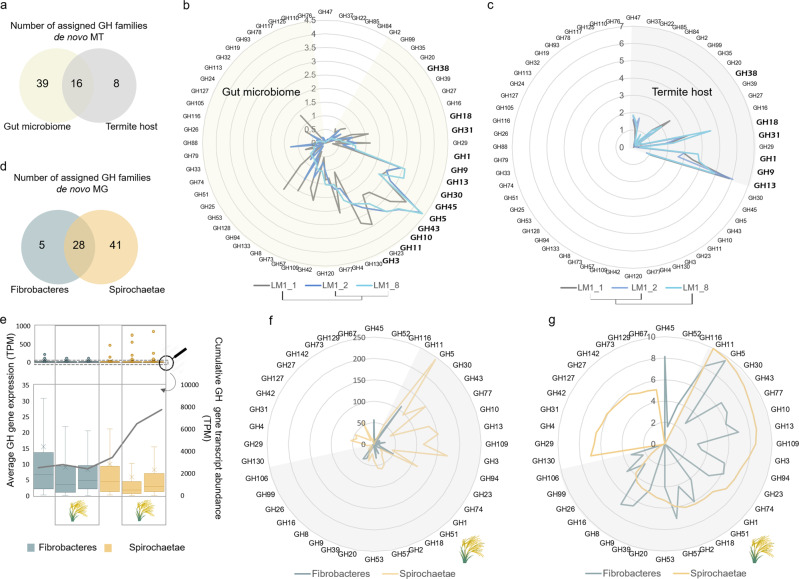
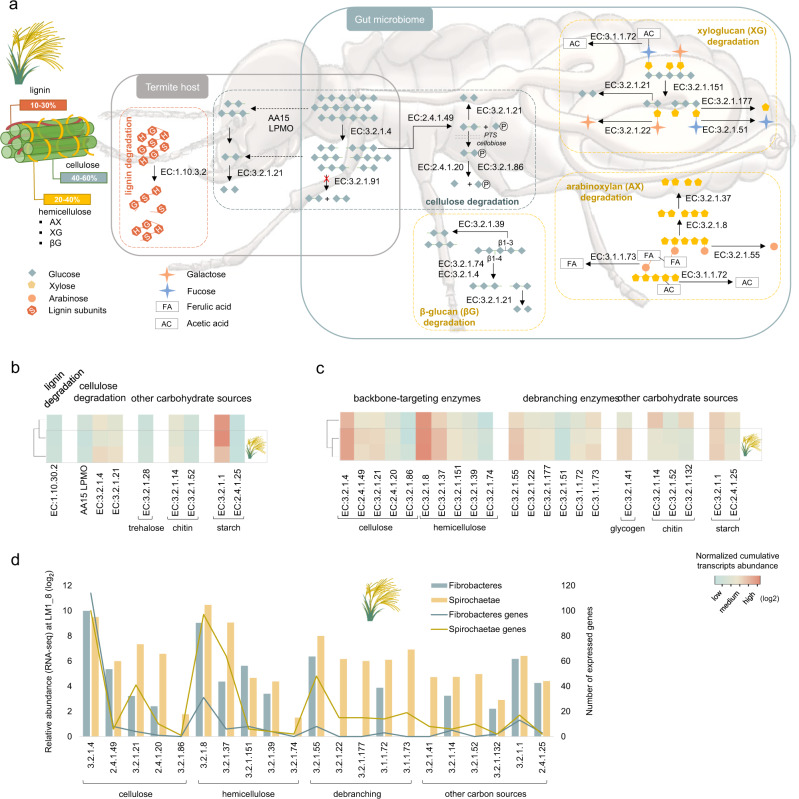
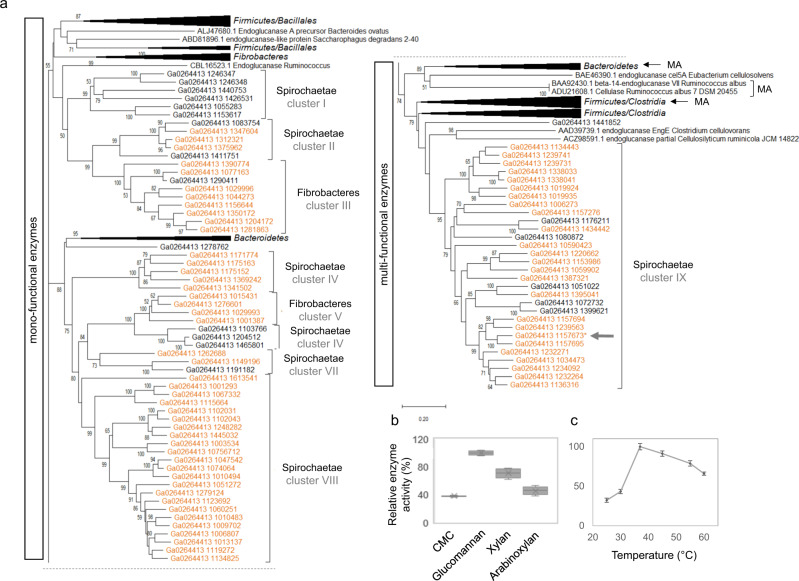
Miscanthus sp. biomass could satisfy future biorefinery value chains. However, its use is largely untapped due to high recalcitrance. The termite and its gut microbiome are considered the most efficient lignocellulose degrading system in nature. Here, we investigate at holobiont level the dynamic adaptation of Cortaritermes sp. to imposed Miscanthus diet, with a long-term objective of overcoming lignocellulose recalcitrance. We use an integrative omics approach combined with enzymatic characterisation of carbohydrate active enzymes from termite gut Fibrobacteres and Spirochaetae. Modified gene expression profiles of gut bacteria suggest a shift towards utilisation of cellulose and arabinoxylan, two main components of Miscanthus lignocellulose. Low identity of reconstructed microbial genomes to closely related species supports the hypothesis of a strong phylogenetic relationship between host and its gut microbiome. This study provides a framework for better understanding the complex lignocellulose degradation by the higher termite gut system and paves a road towards its future bioprospecting.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Lewandowski I, Clifton-Brown JC, Scurlock JMO, Huisman W. Miscanthus: European experience with a novel energy crop. Biomass-. Bioenergy. 2000;19:209–227.
-
- Luo H, et al. Total utilization of Miscanthus biomass, lignin and carbohydrates, using Earth abundant nickel catalyst. ACS Sustain. Chem. Eng. 2016;4:2316–2322.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
