

Temperature compensation in a small rhythmic circuit
- PMID: 32484437
- PMCID: PMC7332291
- DOI: 10.7554/eLife.55470
Temperature compensation in a small rhythmic circuit
Abstract
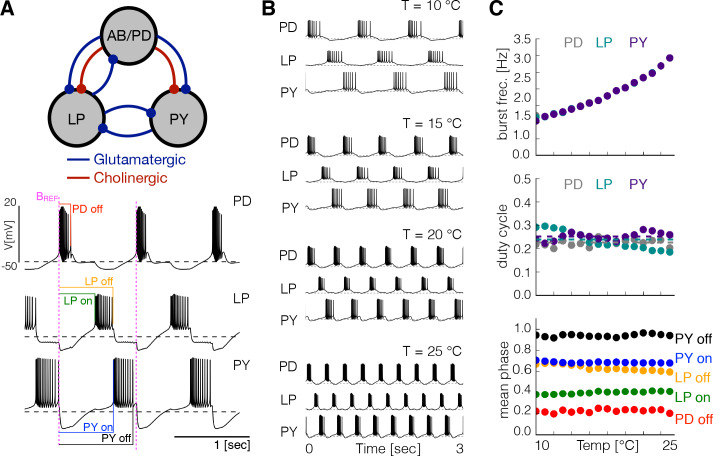
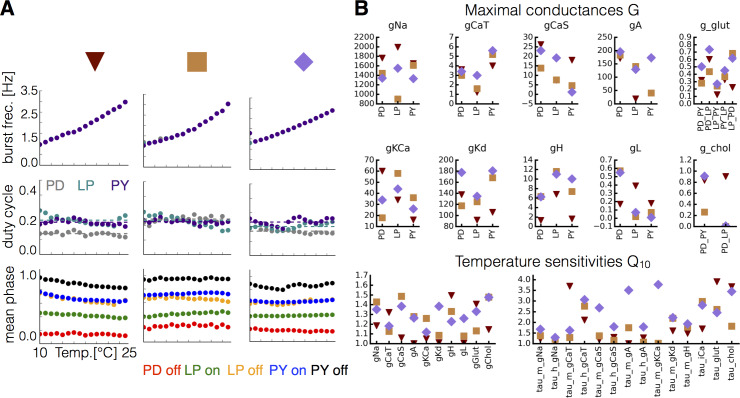
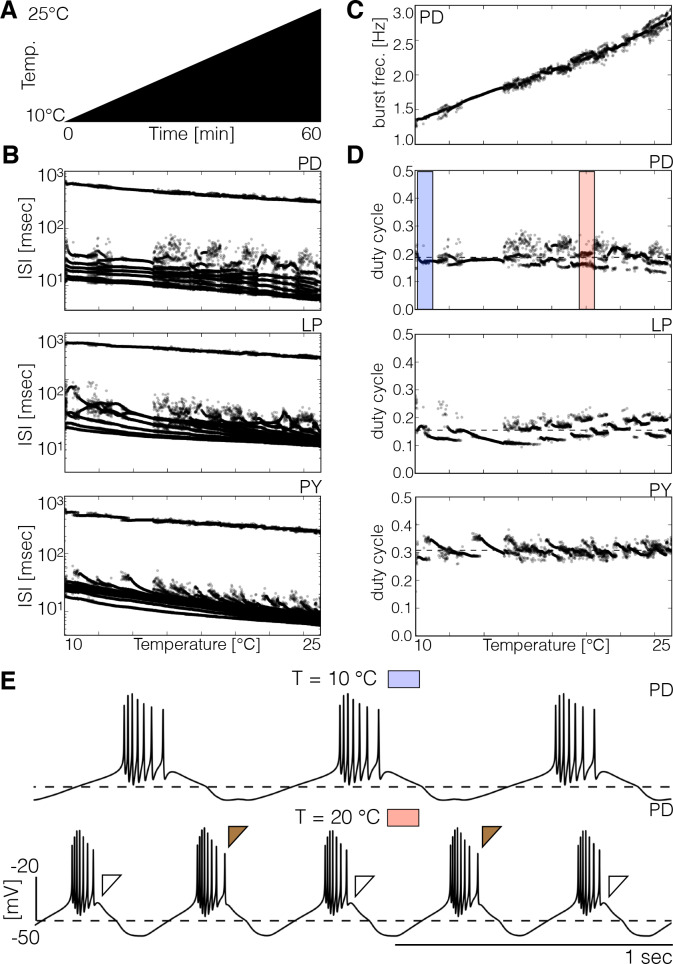
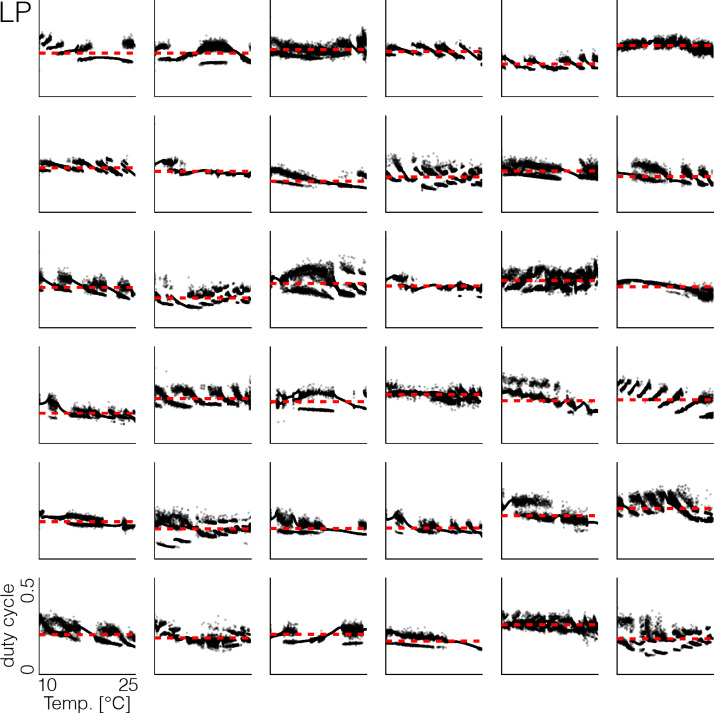
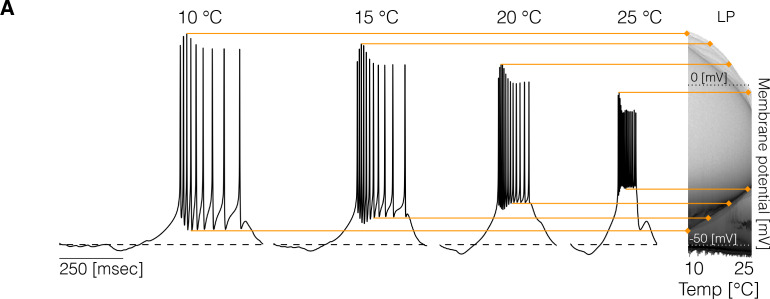
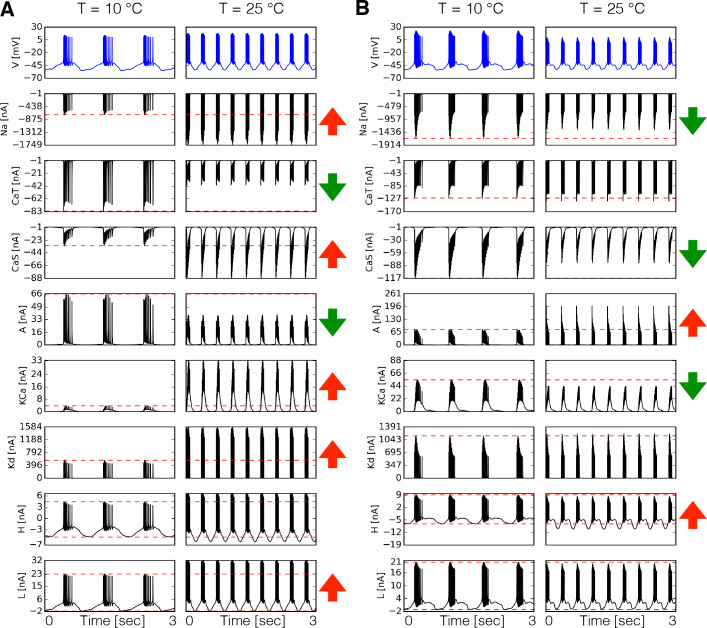
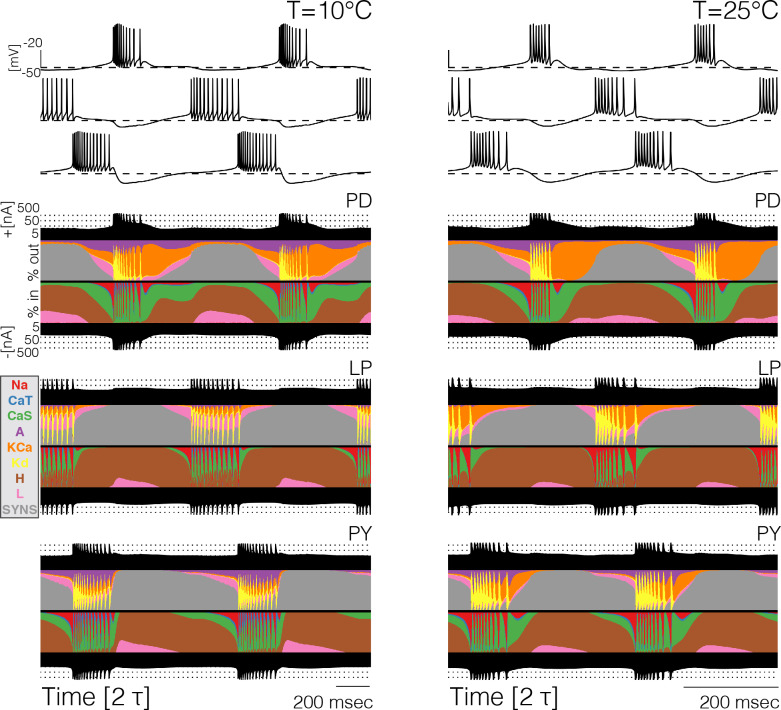
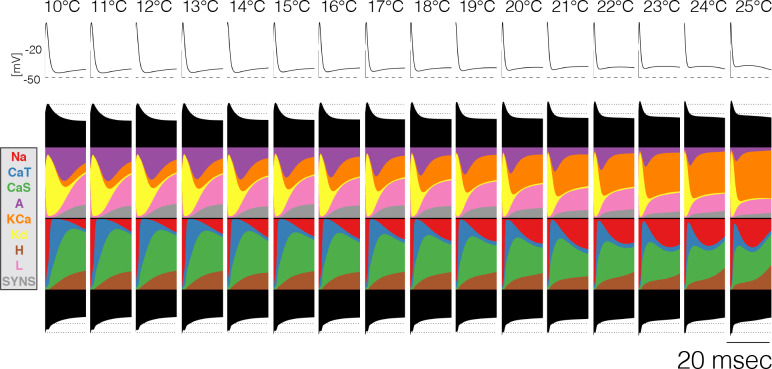
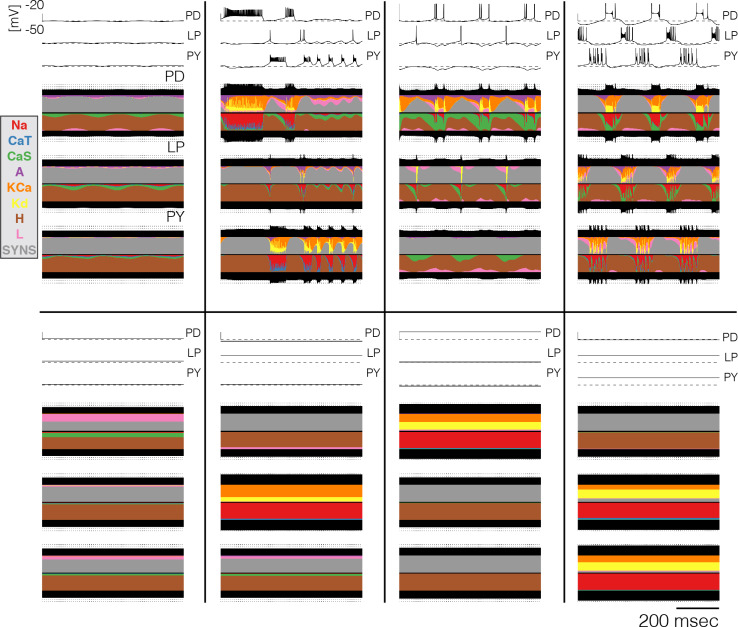
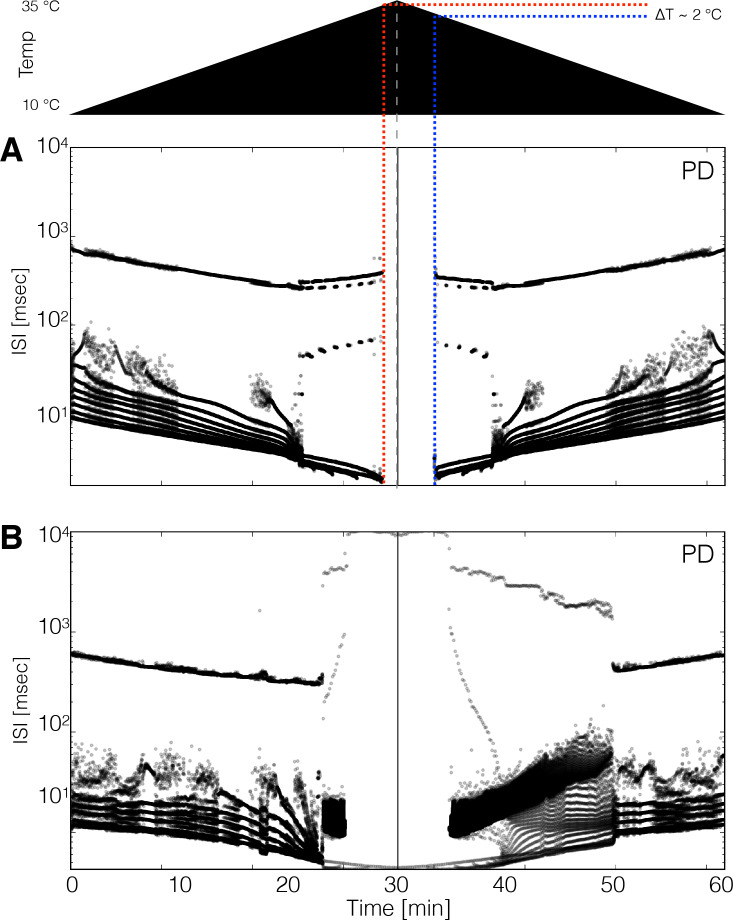
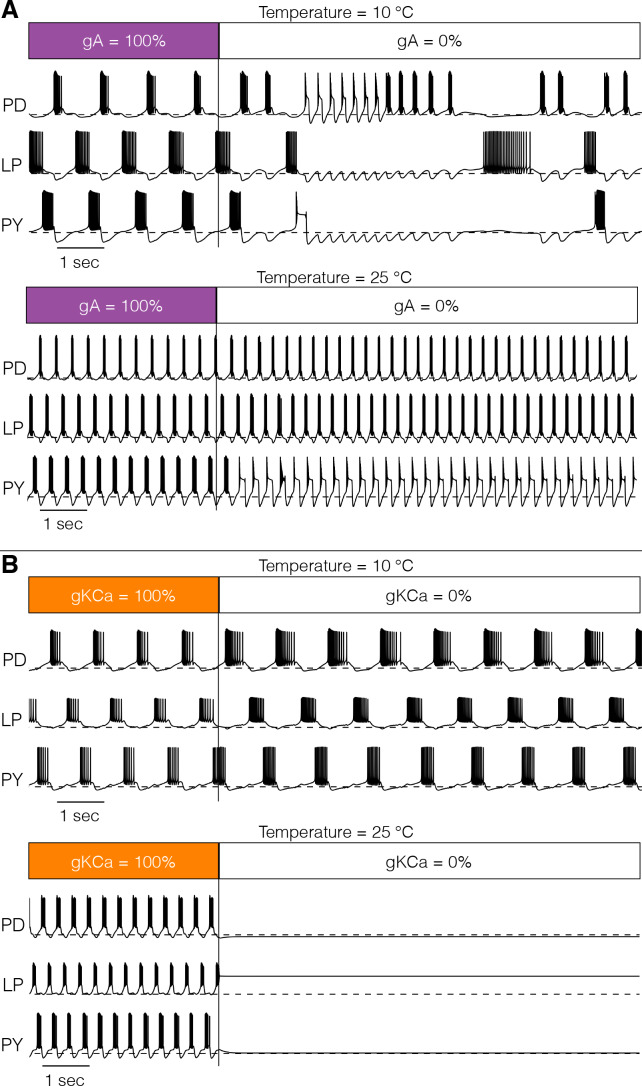
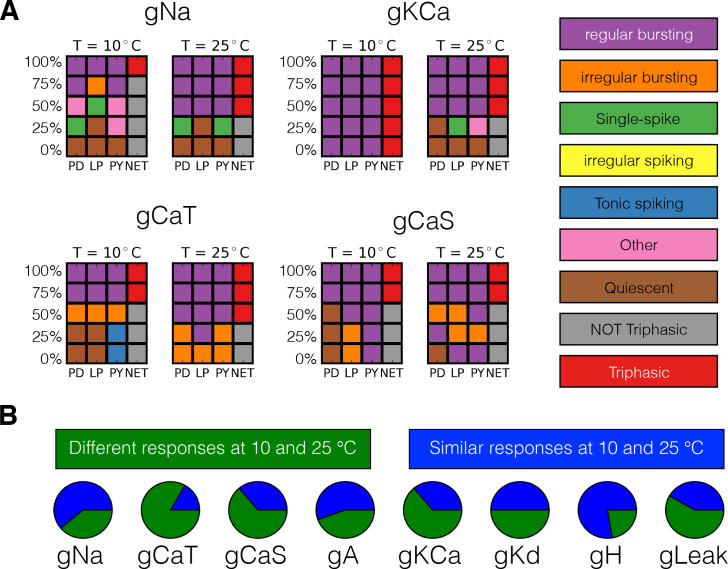
Temperature affects the conductances and kinetics of the ionic channels that underlie neuronal activity. Each membrane conductance has a different characteristic temperature sensitivity, which raises the question of how neurons and neuronal circuits can operate robustly over wide temperature ranges. To address this, we employed computational models of the pyloric network of crabs and lobsters. We produced multiple different models that exhibit a triphasic pyloric rhythm over a range of temperatures and explored the dynamics of their currents and how they change with temperature. Temperature can produce smooth changes in the relative contributions of the currents to neural activity so that neurons and networks undergo graceful transitions in the mechanisms that give rise to their activity patterns. Moreover, responses of the models to deletions of a current can be different at high and low temperatures, indicating that even a well-defined genetic or pharmacological manipulation may produce qualitatively distinct effects depending on the temperature.
Keywords: Q10; ion channels; neuronal oscillators; neuroscience; none; temperature.
© 2020, Alonso and Marder.
Conflict of interest statement
LA, EM No competing interests declared
Figures



References
Publication types
MeSH terms
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
