

ERK2 Phosphorylates PFAS to Mediate Posttranslational Control of De Novo Purine Synthesis
- PMID: 32485148
- PMCID: PMC7306006
- DOI: 10.1016/j.molcel.2020.05.001
ERK2 Phosphorylates PFAS to Mediate Posttranslational Control of De Novo Purine Synthesis
Abstract
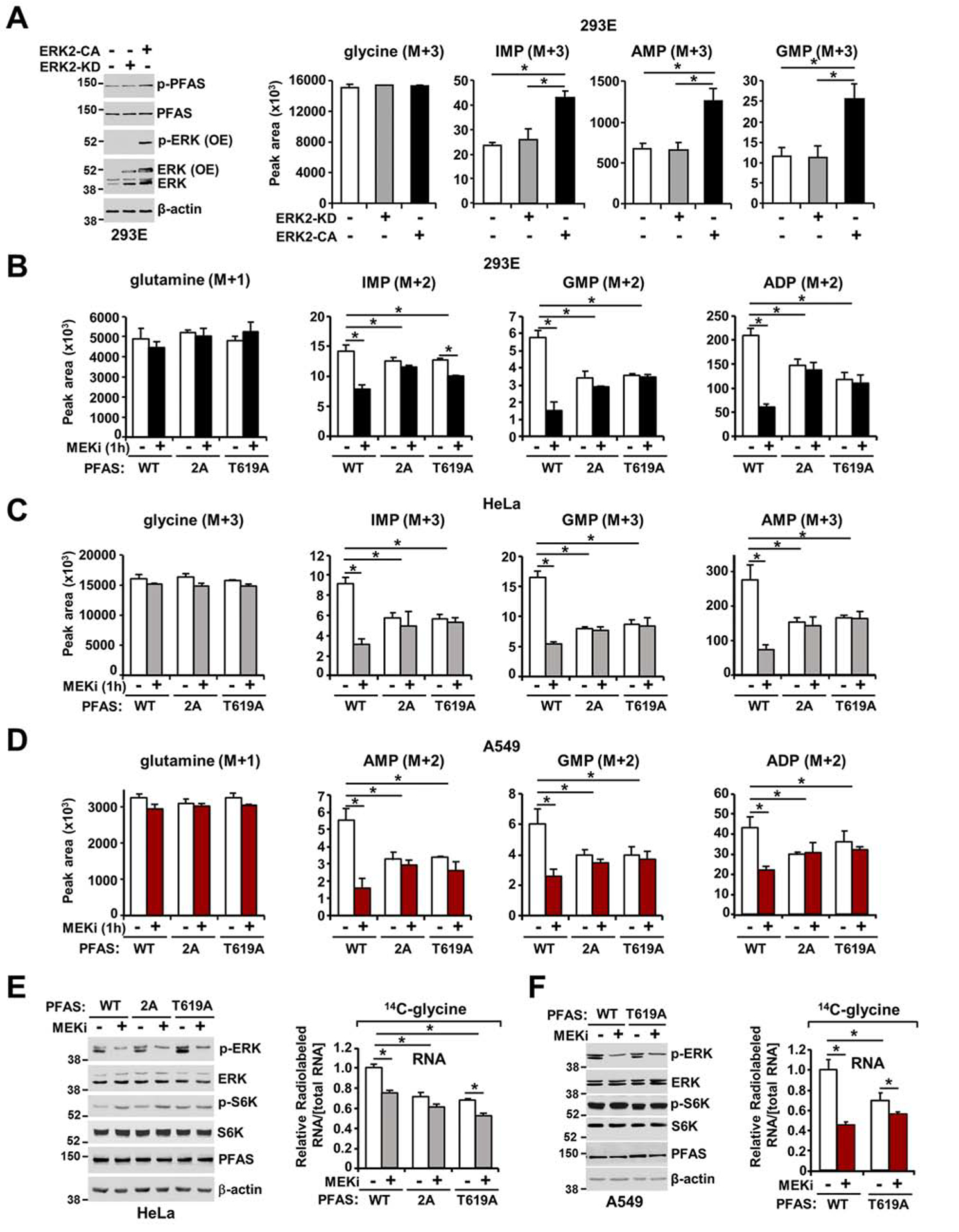
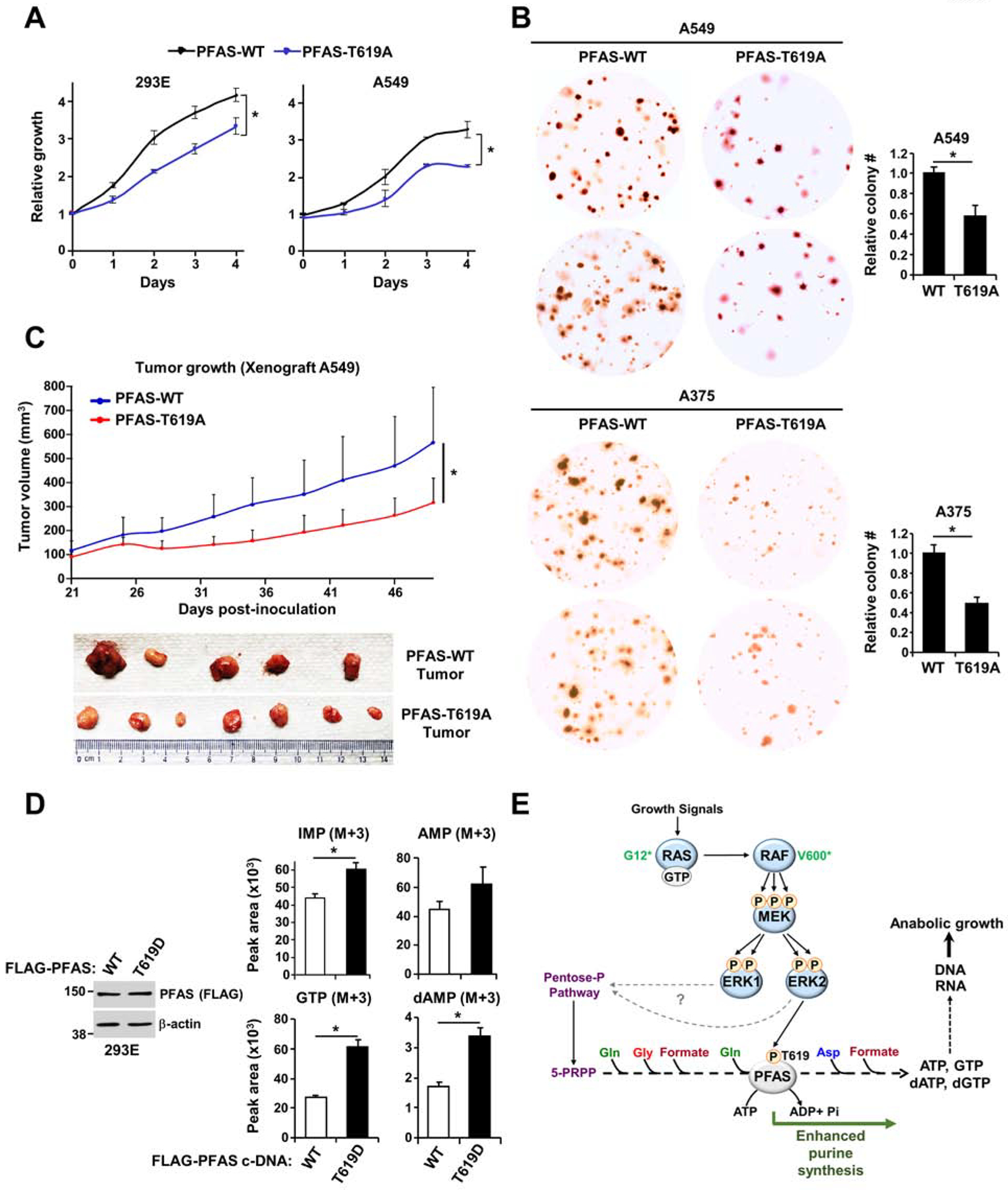
The RAS-ERK/MAPK (RAS-extracellular signal-regulated kinase/mitogen-activated protein kinase) pathway integrates growth-promoting signals to stimulate cell growth and proliferation, at least in part, through alterations in metabolic gene expression. However, examples of direct and rapid regulation of the metabolic pathways by the RAS-ERK pathway remain elusive. We find that physiological and oncogenic ERK signaling activation leads to acute metabolic flux stimulation through the de novo purine synthesis pathway, thereby increasing building block availability for RNA and DNA synthesis, which is required for cell growth and proliferation. We demonstrate that ERK2, but not ERK1, phosphorylates the purine synthesis enzyme PFAS (phosphoribosylformylglycinamidine synthase) at T619 in cells to stimulate de novo purine synthesis. The expression of nonphosphorylatable PFAS (T619A) decreases purine synthesis, RAS-dependent cancer cell-colony formation, and tumor growth. Thus, ERK2-mediated PFAS phosphorylation facilitates the increase in nucleic acid synthesis required for anabolic cell growth and proliferation.
Keywords: ERK; FGAM; MAPK; PFAS; RAS; cancer; nucleotide synthesis; posttranslational modification; purine metabolism; tumor growth.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
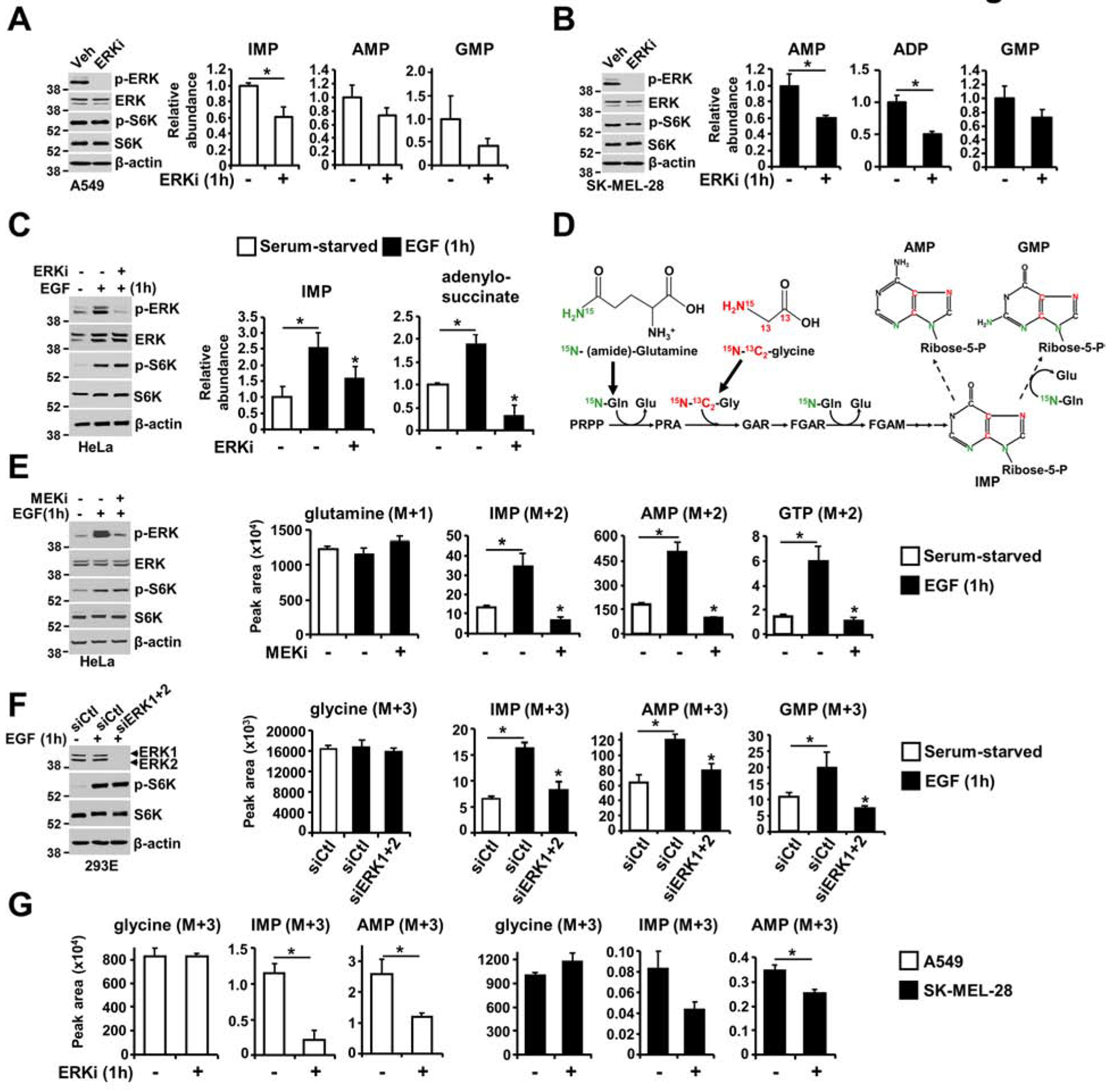
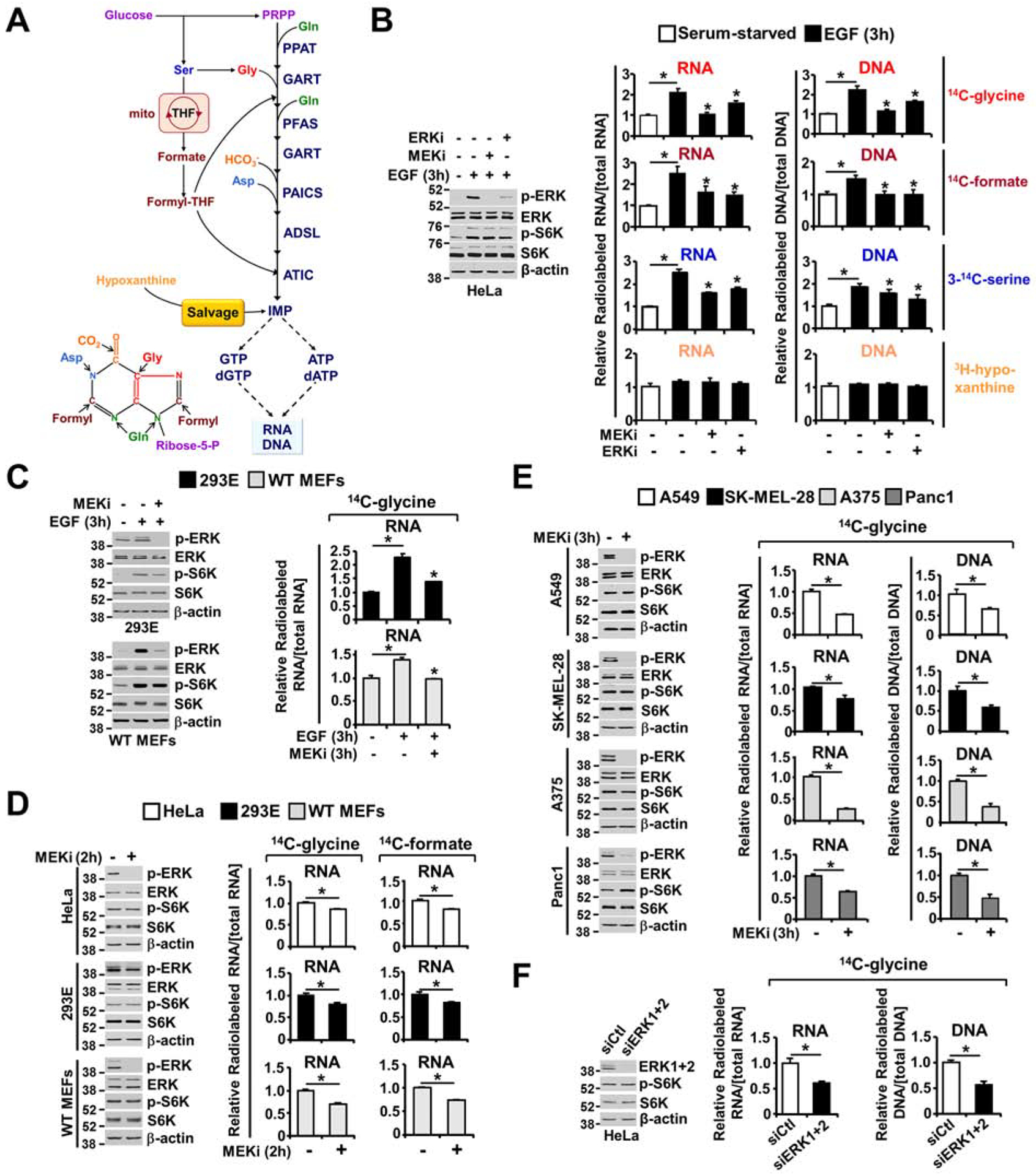
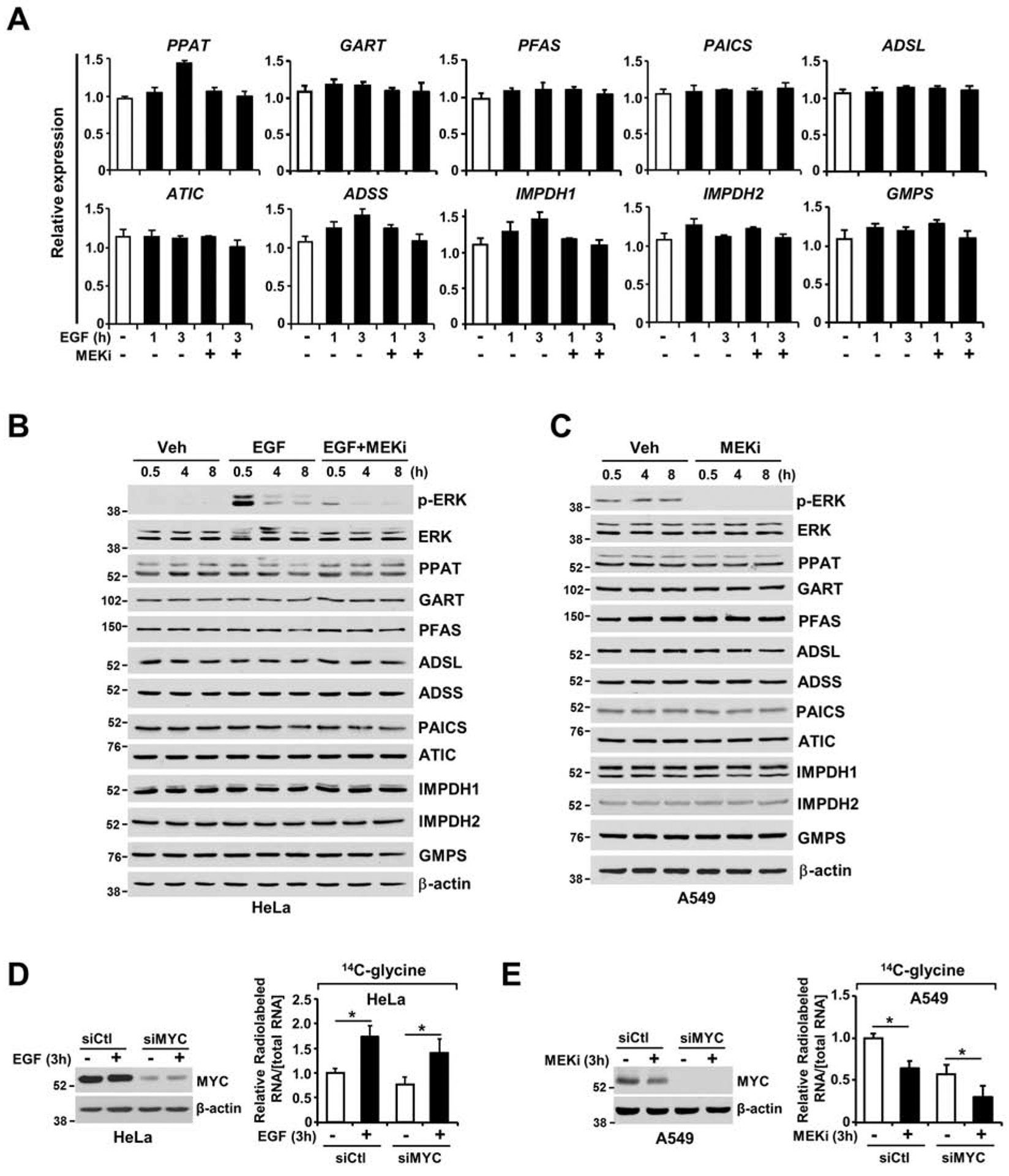
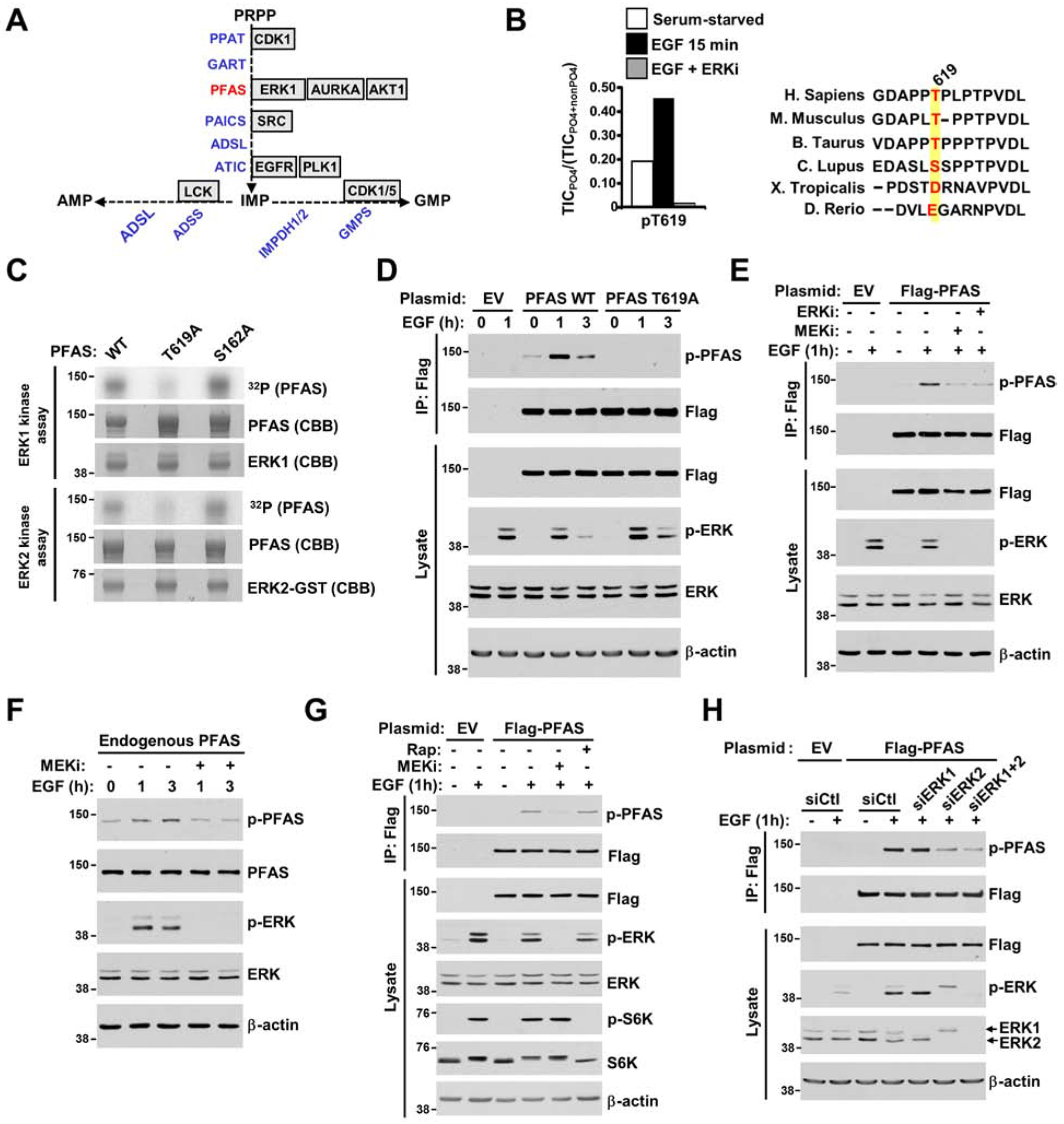
References
-
- Bodoky G, Timcheva C, Spigel DR, La Stella PJ, Ciuleanu TE, Pover G, and Tebbutt NC (2012). A phase II open-label randomized study to assess the efficacy and safety of selumetinib (AZD6244 [ARRY-142886]) versus capecitabine in patients with advanced or metastatic pancreatic cancer who have failed first-line gemcitabine therapy. Investigational new drugs 30, 1216–1223. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
