

The Procoagulant Snake Venom Serine Protease Potentially Having a Dual, Blood Coagulation Factor V and X-Activating Activity
- PMID: 32485989
- PMCID: PMC7354534
- DOI: 10.3390/toxins12060358
The Procoagulant Snake Venom Serine Protease Potentially Having a Dual, Blood Coagulation Factor V and X-Activating Activity
Abstract
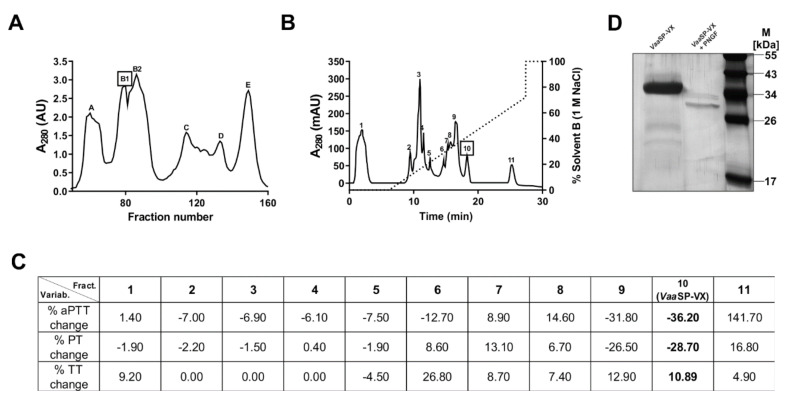
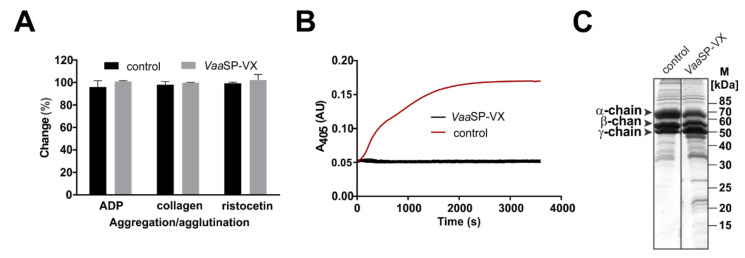
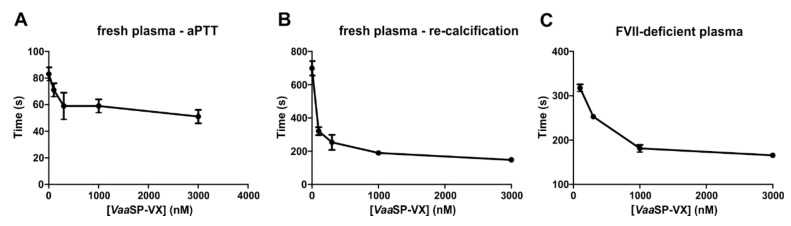
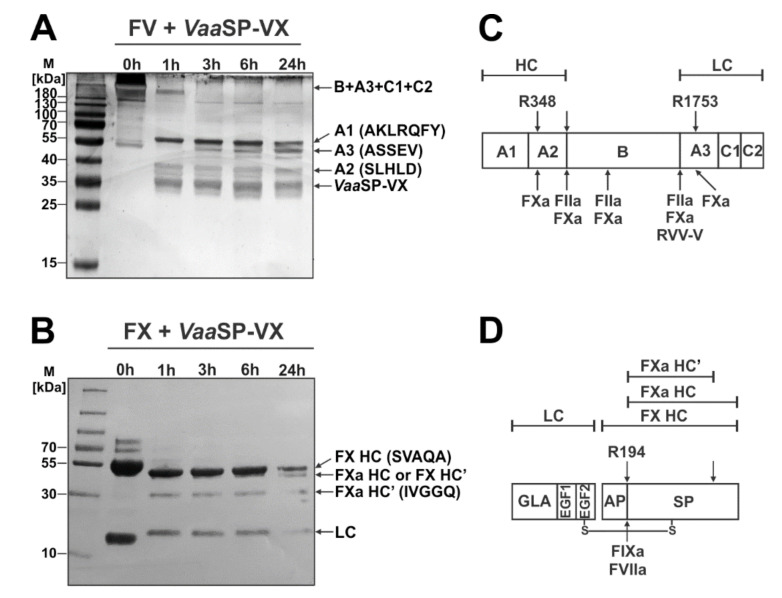
A procoagulant snake venom serine protease was isolated from the venom of the nose-horned viper (Vipera ammodytes ammodytes). This 34 kDa glycoprotein, termed VaaSP-VX, possesses five kDa N-linked carbohydrates. Amino acid sequencing showed VaaSP-VX to be a chymotrypsin-like serine protease. Structurally, it is highly homologous to VaaSP-6 from the same venom and to nikobin from the venom of Vipera nikolskii, neither of which have known functions. VaaSP-VX does not affect platelets. The specific proteolysis of blood coagulation factors X and V by VaaSP-VX suggests that its blood-coagulation-inducing effect is due to its ability to activate these two blood coagulation factors, which following activation, combine to form the prothrombinase complex. VaaSP-VX may thus represent the first example of a serine protease with such a dual activity, which makes it a highly suitable candidate to replace diluted Russell's viper venom in lupus anticoagulant testing, thus achieving greater reliability of the analysis. As a blood-coagulation-promoting substance that is resistant to serpin inhibition, VaaSP-VX is also interesting from the therapeutic point of view for treating patients suffering from hemophilia.
Keywords: FV activator; FX activator; procoagulant; serine protease; snake venom.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
Similar articles
-
The pro-coagulant fibrinogenolytic serine protease isoenzymes purified from Daboia russelii russelii venom coagulate the blood through factor V activation: role of glycosylation on enzymatic activity.PLoS One. 2014 Feb 10;9(2):e86823. doi: 10.1371/journal.pone.0086823. eCollection 2014. PLoS One. 2014. PMID: 24520323 Free PMC article.
-
Comparative study of anticoagulant and procoagulant properties of 28 snake venoms from families Elapidae, Viperidae, and purified Russell's viper venom-factor X activator (RVV-X).Toxicon. 2010 Sep 15;56(4):544-53. doi: 10.1016/j.toxicon.2010.05.012. Toxicon. 2010. PMID: 20677373 Free PMC article.
-
VaSP1, catalytically active serine proteinase from Vipera ammodytes ammodytes venom with unconventional active site triad.Toxicon. 2014 Jan;77:93-104. doi: 10.1016/j.toxicon.2013.11.007. Epub 2013 Nov 20. Toxicon. 2014. PMID: 24269689
-
Procoagulant proteins from snake venoms.Haemostasis. 2001 May-Dec;31(3-6):218-24. doi: 10.1159/000048066. Haemostasis. 2001. PMID: 11910188 Review.
-
Snake venom activators of factor X: an overview.Haemostasis. 2001 May-Dec;31(3-6):225-33. doi: 10.1159/000048067. Haemostasis. 2001. PMID: 11910189 Review.
Cited by
-
Hump-Nosed Pit Viper (Hypnale hypnale) Venom-Induced Irreversible Red Blood Cell Aggregation, Inhibition by Monovalent Anti-Venom and N-Acetylcysteine.Cells. 2024 Jun 7;13(12):994. doi: 10.3390/cells13120994. Cells. 2024. PMID: 38920625 Free PMC article.
-
Tiny but Mighty: Vipera ammodytes meridionalis (Eastern Long-Nosed Viper) Ontogenetic Venom Variations in Procoagulant Potency and the Impact on Antivenom Efficacies.Toxins (Basel). 2024 Sep 14;16(9):396. doi: 10.3390/toxins16090396. Toxins (Basel). 2024. PMID: 39330854 Free PMC article.
-
Snake Venom Compounds: A New Frontier in the Battle Against Antibiotic-Resistant Infections.Toxins (Basel). 2025 May 1;17(5):221. doi: 10.3390/toxins17050221. Toxins (Basel). 2025. PMID: 40423304 Free PMC article. Review.
-
Metabolomics and proteomics: synergistic tools for understanding snake venom inhibition.Arch Toxicol. 2025 Mar;99(3):915-934. doi: 10.1007/s00204-024-03947-4. Epub 2025 Jan 6. Arch Toxicol. 2025. PMID: 39760869 Review.
-
Heating up the Blunts: Prothrombin Activation, with Factor Va as an Obligate Cofactor, Is the Dominant Procoagulant Mechanism of Blunt-Nosed Viper Venoms (Macrovipera Species).Toxins (Basel). 2025 Aug 8;17(8):398. doi: 10.3390/toxins17080398. Toxins (Basel). 2025. PMID: 40864074 Free PMC article.
References
-
- Alvarez-Flores M.P., Faria F., de Andrade S.A., Chudzinski-Tavassi A.M. Snake venom components affecting the coagulation system. In: Gopalakrishnakone P., Inagaki H., Vogel C.W., Mukherjee A.K., Rahmy T.R., editors. Snake Venoms. Springer; Dordrecht, The Netherlands: 2017. pp. 417–436.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
