

TNF-alpha-induced microglia activation requires miR-342: impact on NF-kB signaling and neurotoxicity
- PMID: 32488063
- PMCID: PMC7265562
- DOI: 10.1038/s41419-020-2626-6
TNF-alpha-induced microglia activation requires miR-342: impact on NF-kB signaling and neurotoxicity
Abstract
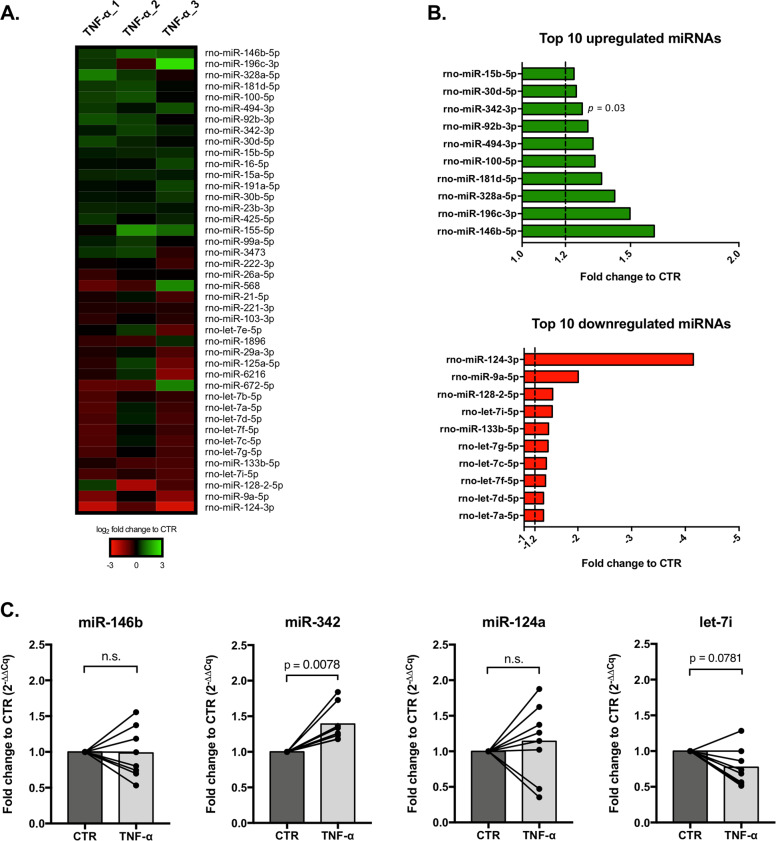
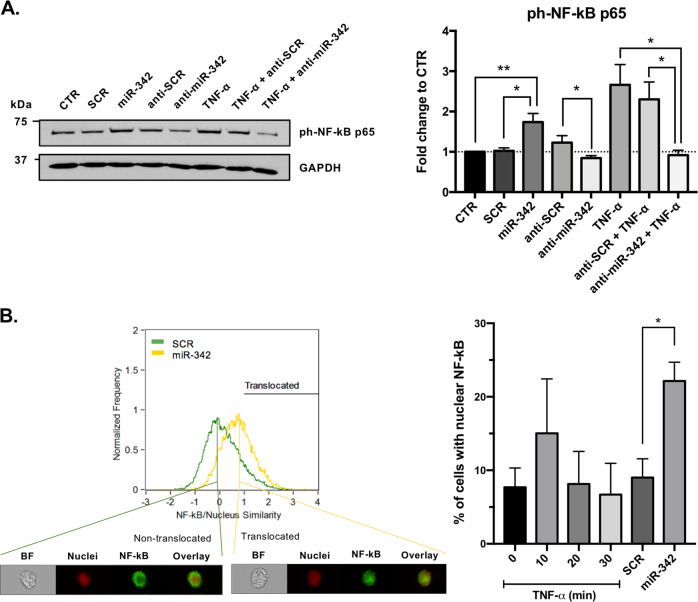
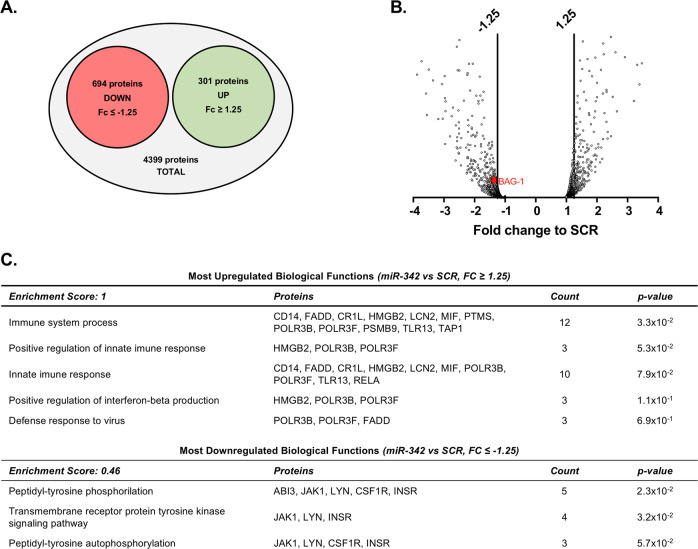
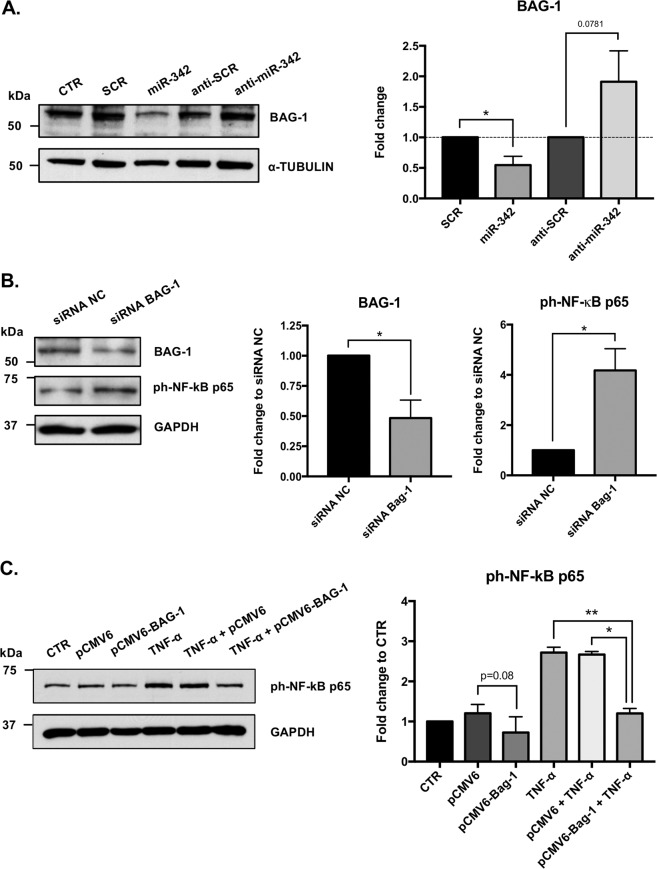
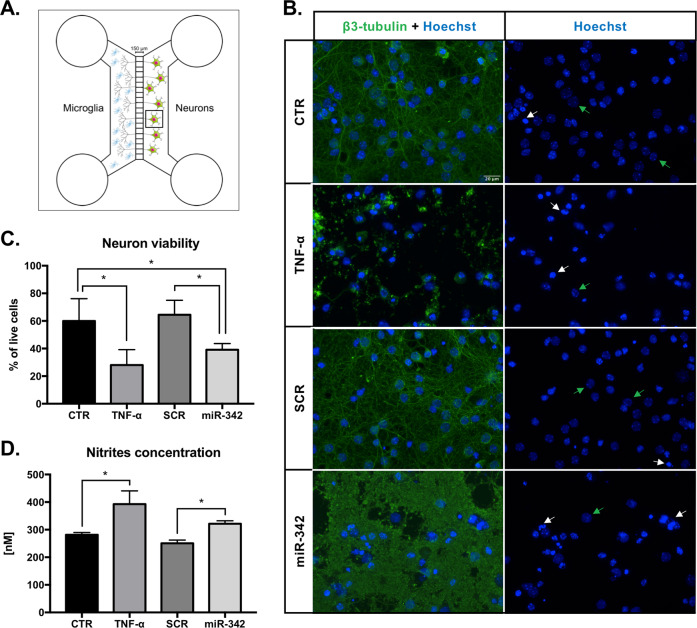
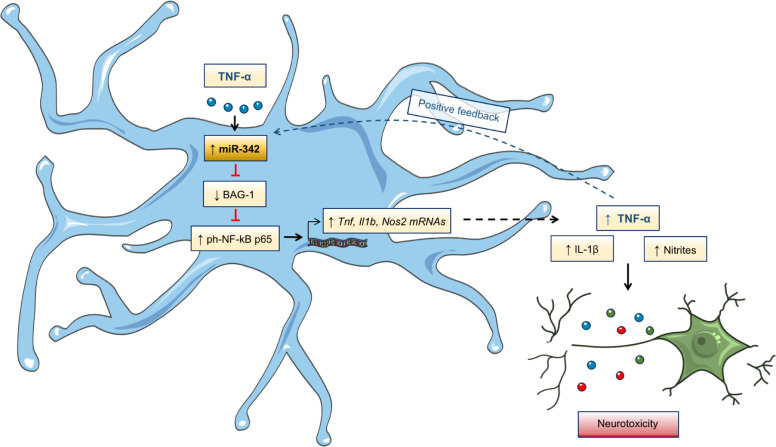
Growing evidences suggest that sustained neuroinflammation, caused by microglia overactivation, is implicated in the development and aggravation of several neurological and psychiatric disorders. In some pathological conditions, microglia produce increased levels of cytotoxic and inflammatory mediators, such as tumor necrosis factor alpha (TNF-α), which can reactivate microglia in a positive feedback mechanism. However, specific molecular mediators that can be effectively targeted to control TNF-α-mediated microglia overactivation, are yet to be uncovered. In this context, we aim to identify novel TNF-α-mediated micro(mi)RNAs and to dissect their roles in microglia activation, as well as to explore their impact on the cellular communication with neurons. A miRNA microarray, followed by RT-qPCR validation, was performed on TNF-α-stimulated primary rat microglia. Gain- and loss-of-function in vitro assays and proteomic analysis were used to dissect the role of miR-342 in microglia activation. Co-cultures of microglia with hippocampal neurons, using a microfluidic system, were performed to understand the impact on neurotoxicity. Stimulation of primary rat microglia with TNF-α led to an upregulation of Nos2, Tnf, and Il1b mRNAs. In addition, ph-NF-kB p65 levels were also increased. miRNA microarray analysis followed by RT-qPCR validation revealed that TNF-α stimulation induced the upregulation of miR-342. Interestingly, miR-342 overexpression in N9 microglia was sufficient to activate the NF-kB pathway by inhibiting BAG-1, leading to increased secretion of TNF-α and IL-1β. Conversely, miR-342 inhibition led to a strong decrease in the levels of these cytokines after TNF-α activation. In fact, both TNF-α-stimulated and miR-342-overexpressing microglia drastically affected neuron viability. Remarkably, increased levels of nitrites were detected in the supernatants of these co-cultures. Globally, our findings show that miR-342 is a crucial mediator of TNF-α-mediated microglia activation and a potential target to tackle microglia-driven neuroinflammation.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures


References
-
- Yirmiya R, Rimmerman N, Reshef R. Depression as a microglial disease. Trends Neurosci. 2015;38:637–658. - PubMed
-
- Thion MS, Ginhoux F, Garel S. Microglia and early brain development: an intimate journey. Science. 2018;362:185–189. - PubMed
-
- Ransohoff RM, Perry VH. Microglial physiology: unique stimuli, specialized responses. Annu. Rev. Immunol. 2009;27:119–145. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
