

Expression profiles and function of IL6 in polymorphonuclear myeloid-derived suppressor cells
- PMID: 32488308
- PMCID: PMC11027602
- DOI: 10.1007/s00262-020-02620-w
Expression profiles and function of IL6 in polymorphonuclear myeloid-derived suppressor cells
Abstract
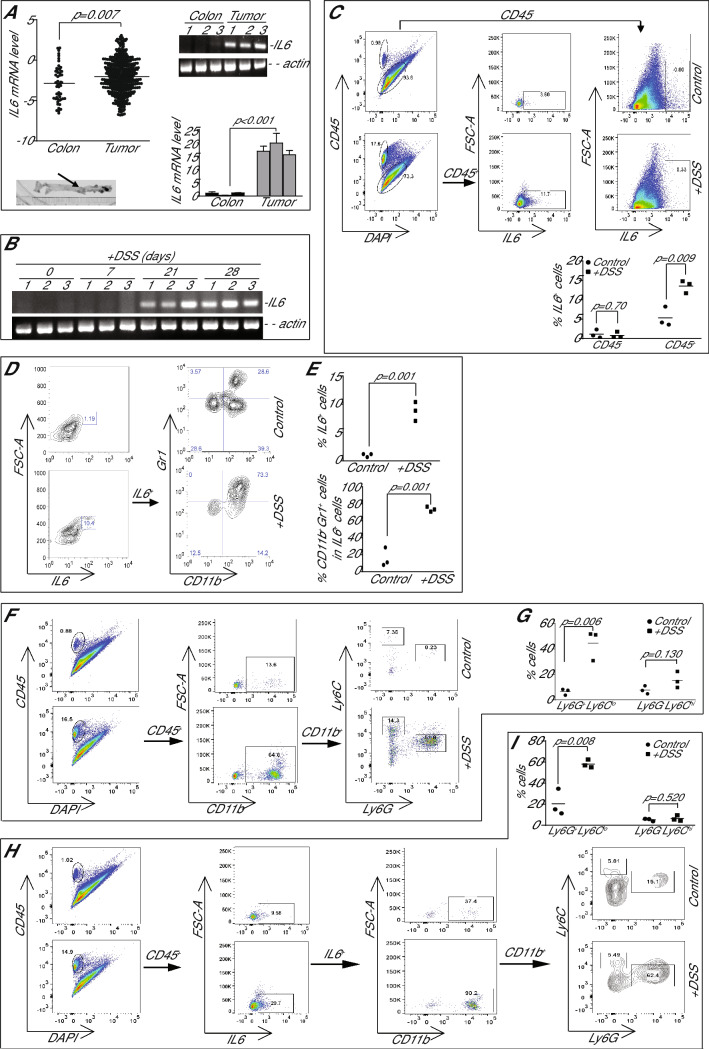
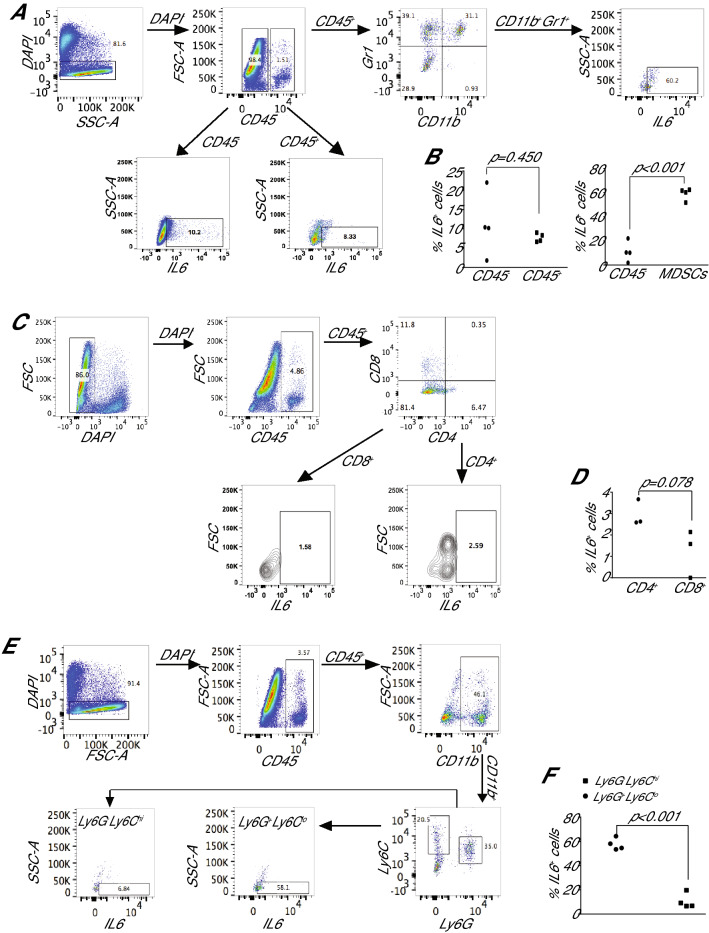
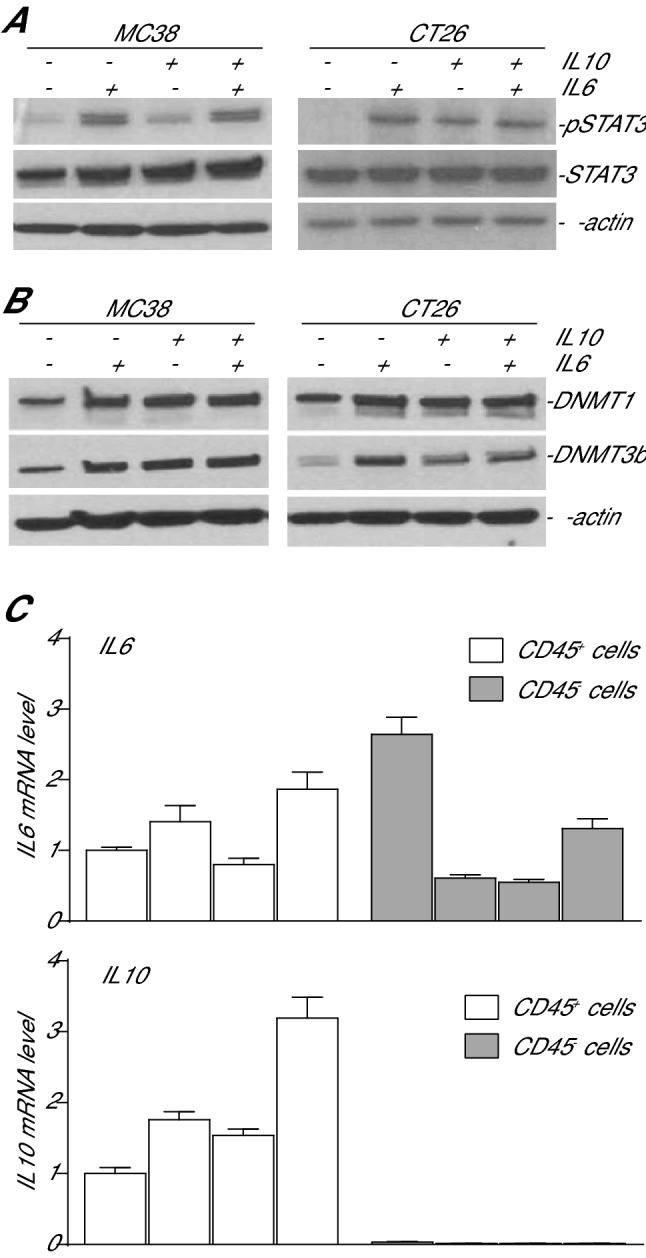
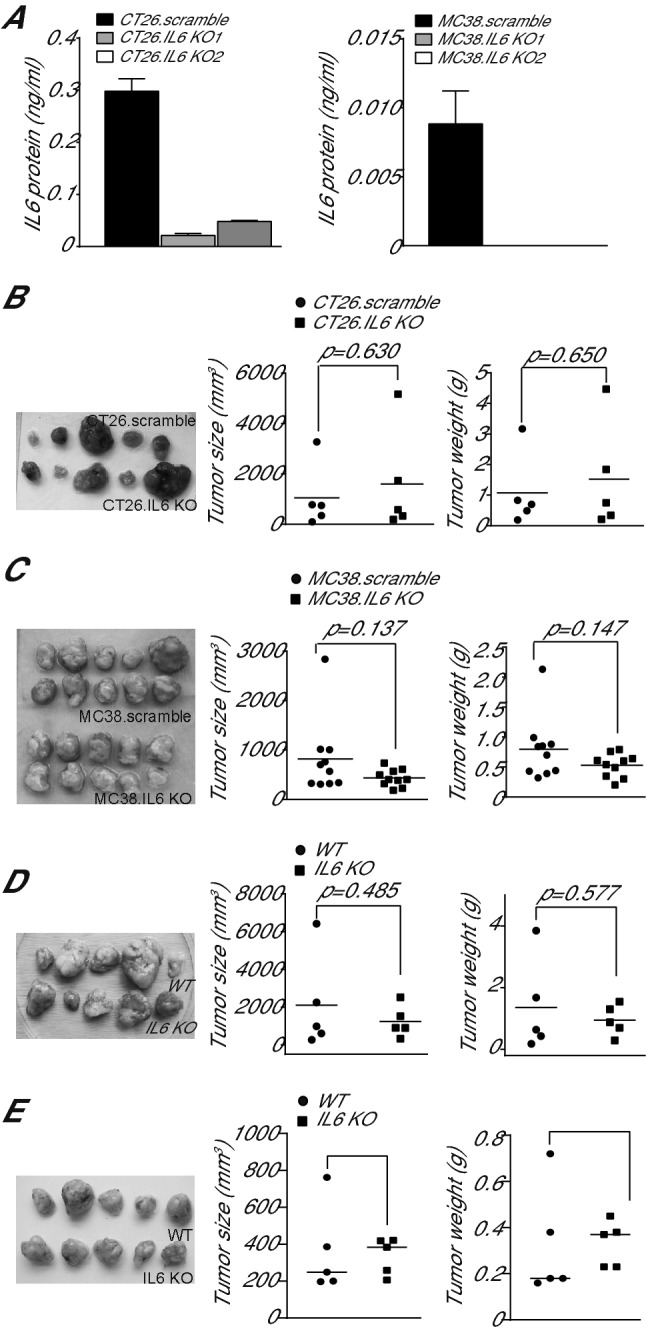
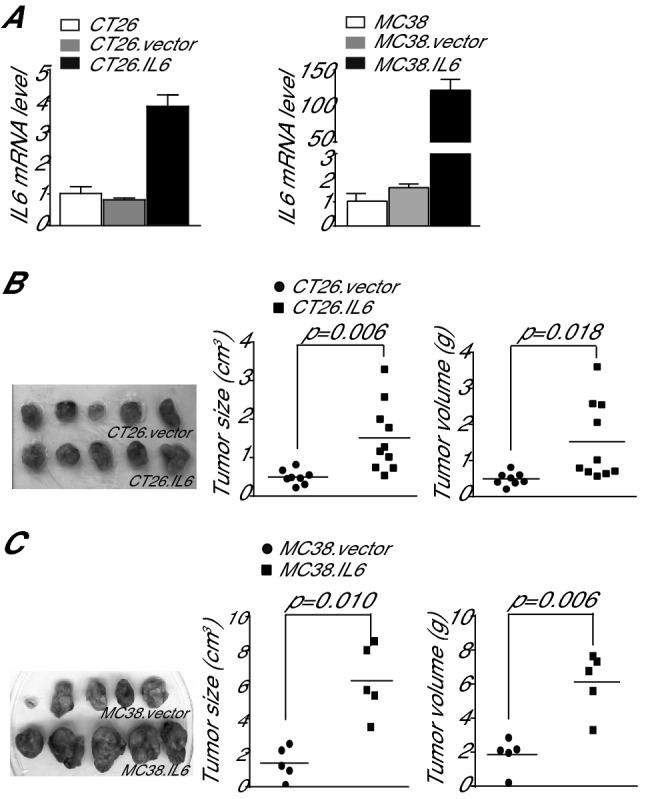
IL6 is an inflammatory cytokine with pleiotropic functions in both immune and nonimmune cells, and its expression level is inversely correlated with disease prognosis in patients with cancer. However, blocking IL6 alone has only yielded minimal efficacy in human cancer patients. We aimed at defining IL6 expression profiles under inflammatory conditions and cancer, and elucidating the mechanism underlying IL6 intrinsic signaling in colon carcinoma. We report here that colonic inflammation induces IL6 expression primarily in the CD11b+Ly6G+Ly6Clo polymorphonuclear myeloid-derived suppressor cells (PMN-MDSC) in colon. Although both tumor cells, T cells and myeloid cells all express IL6, PMN-MDSCs are the primary cell type that express IL6 in colon carcinoma, suggesting that IL6 up-regulation is a response to inflammation in colon epithelium and tumor microenvironment. Furthermore, we determined that IL6 activates STAT3 to up-regulate DNMT1 and DNMT3b expression in colon tumor cells, thereby revealing an epigenetic mechanism that mediates the IL6-STAT3 signaling pathway in colon carcinoma. Surprisingly, knocking out IL6 in colon tumor cells did not significantly alter tumor growth in WT mice. Conversely, IL6-sufficient colon and pancreatic tumor grow at similar rate in WT and IL6-deficient mice. However, overexpression of IL6 in colon tumor cells significantly increases tumor growth in vivo. Our findings determine that a high tumor local IL6 threshold is essential for IL6 function in colon tumor promotion and targeting the IL6-expressing PMN-MDSCs is potentially an effective approach to suppress colon tumor growth in vivo.
Keywords: Colon tumor; Colonic inflammation; DNMT; IL6; MDSCs; STAT3.
Conflict of interest statement
The authors have declared that no conflict of interest exists.
Figures

References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
