

Synaptotagmin-1 is the Ca2+ sensor for fast striatal dopamine release
- PMID: 32490813
- PMCID: PMC7319770
- DOI: 10.7554/eLife.58359
Synaptotagmin-1 is the Ca2+ sensor for fast striatal dopamine release
Abstract
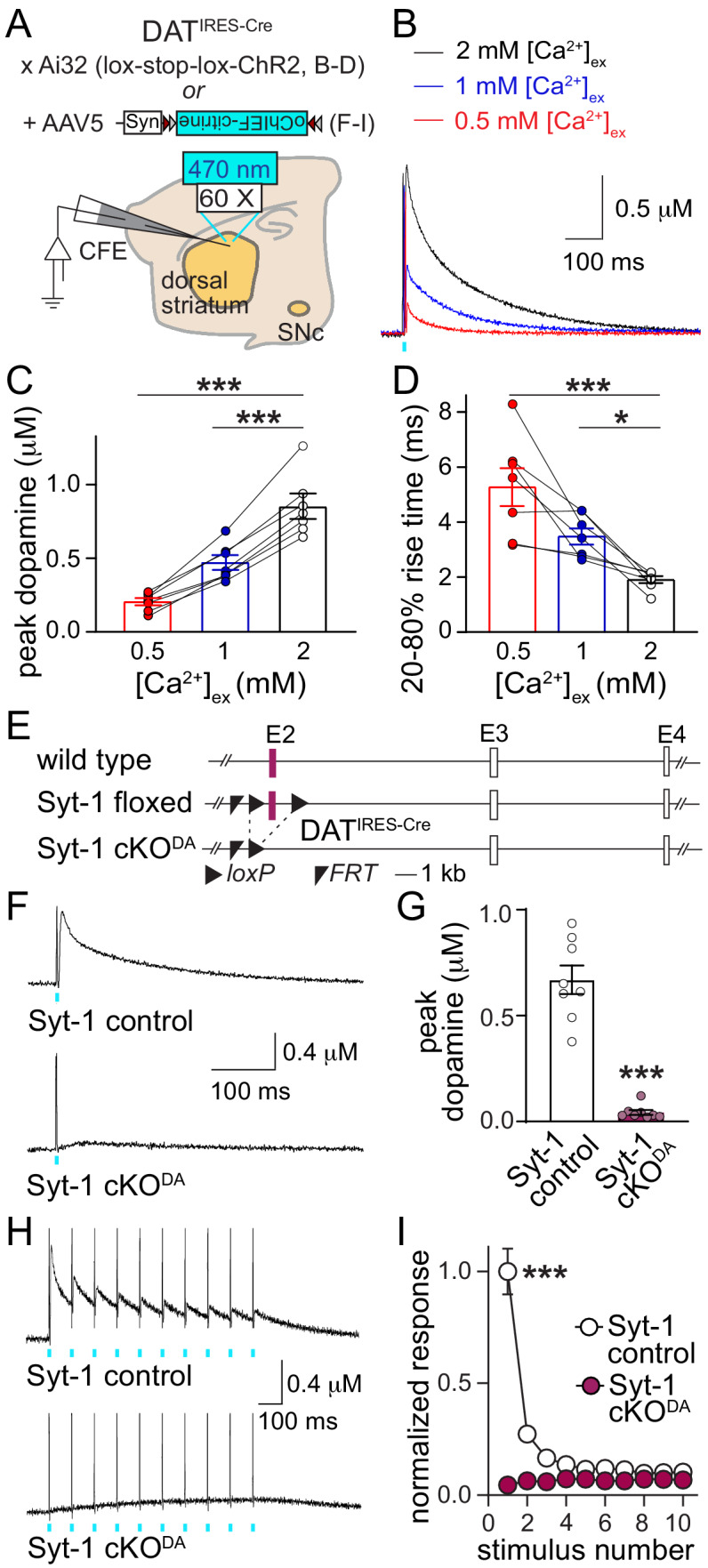
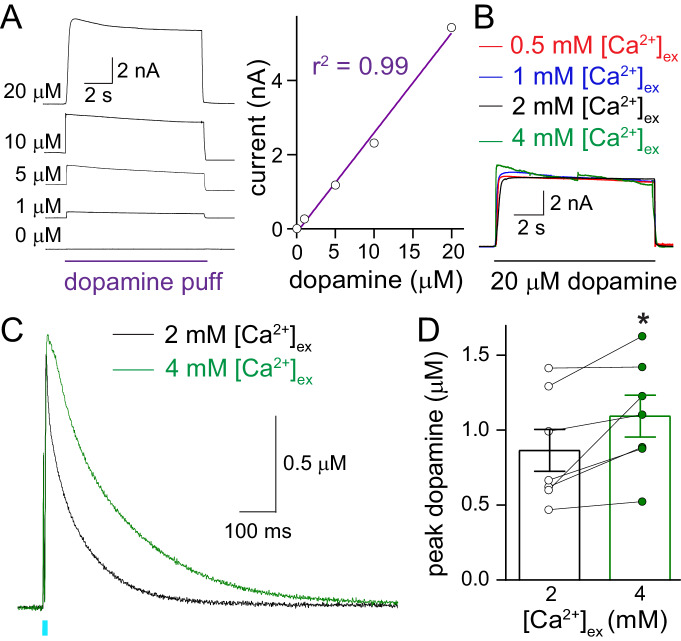
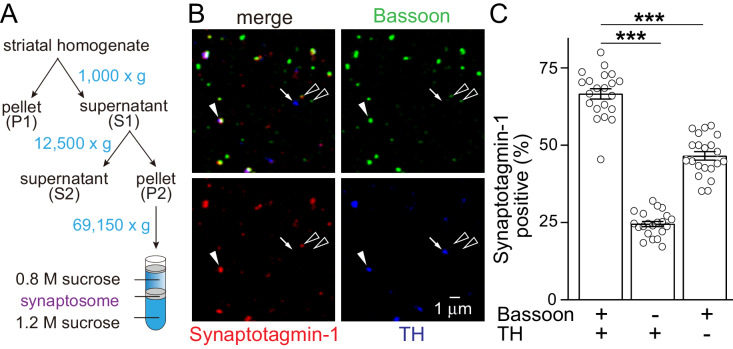
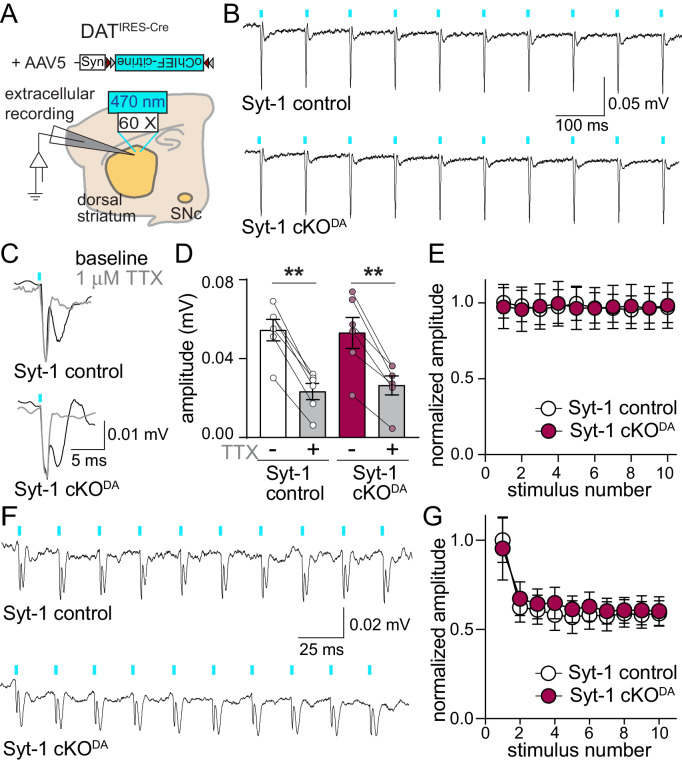
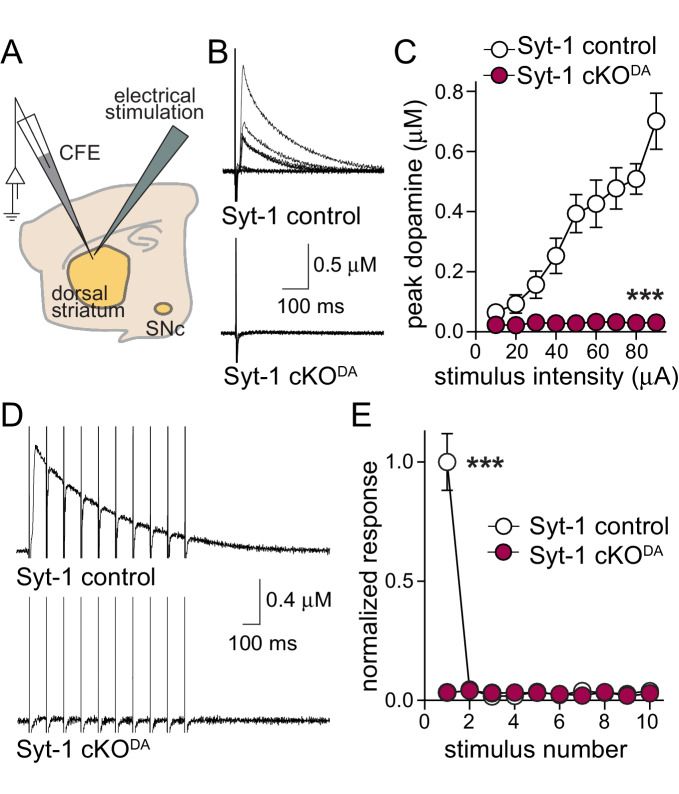
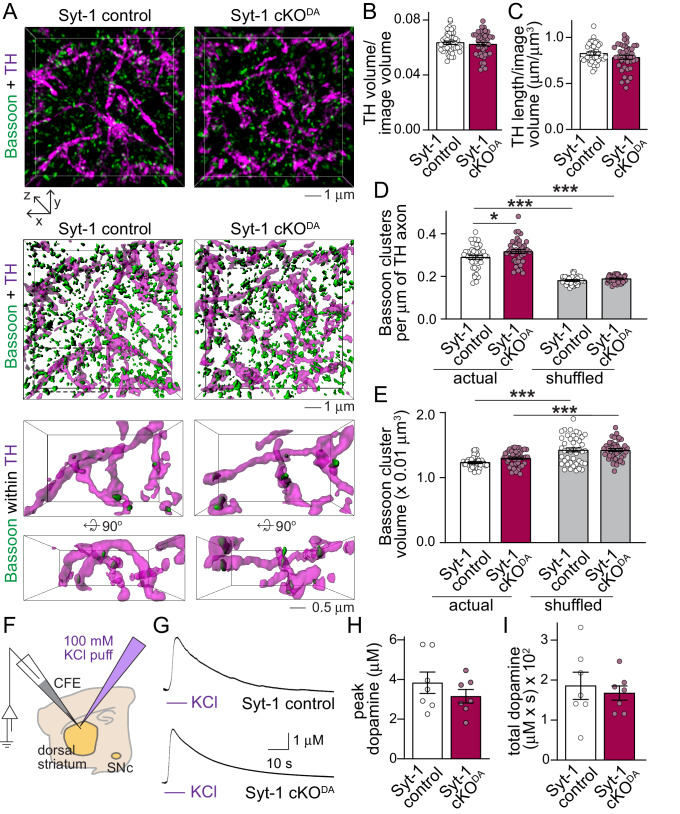
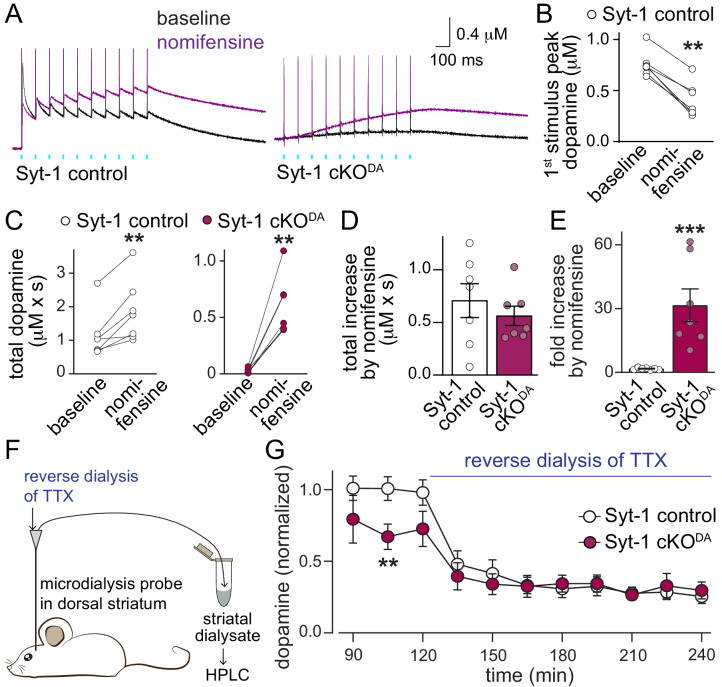
Dopamine powerfully controls neural circuits through neuromodulation. In the vertebrate striatum, dopamine adjusts cellular functions to regulate behaviors across broad time scales, but how the dopamine secretory system is built to support fast and slow neuromodulation is not known. Here, we set out to identify Ca2+-triggering mechanisms for dopamine release. We find that synchronous dopamine secretion is abolished in acute brain slices of conditional knockout mice in which Synaptotagmin-1 is removed from dopamine neurons. This indicates that Synaptotagmin-1 is the Ca2+ sensor for fast dopamine release. Remarkably, dopamine release induced by strong depolarization and asynchronous release during stimulus trains are unaffected by Synaptotagmin-1 knockout. Microdialysis further reveals that these modes and action potential-independent release provide significant amounts of extracellular dopamine in vivo. We propose that the molecular machinery for dopamine secretion has evolved to support fast and slow signaling modes, with fast release requiring the Ca2+ sensor Synaptotagmin-1.
Keywords: calcium sensor; cell biology; dopamine; mouse; neuromodulation; neuroscience; secretion; striatum.
© 2020, Banerjee et al.
Conflict of interest statement
AB, JL, PN, CL, PK No competing interests declared
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
