

Assembly and Function of a Bioengineered Human Liver for Transplantation Generated Solely from Induced Pluripotent Stem Cells
- PMID: 32492423
- PMCID: PMC7734598
- DOI: 10.1016/j.celrep.2020.107711
Assembly and Function of a Bioengineered Human Liver for Transplantation Generated Solely from Induced Pluripotent Stem Cells
Abstract
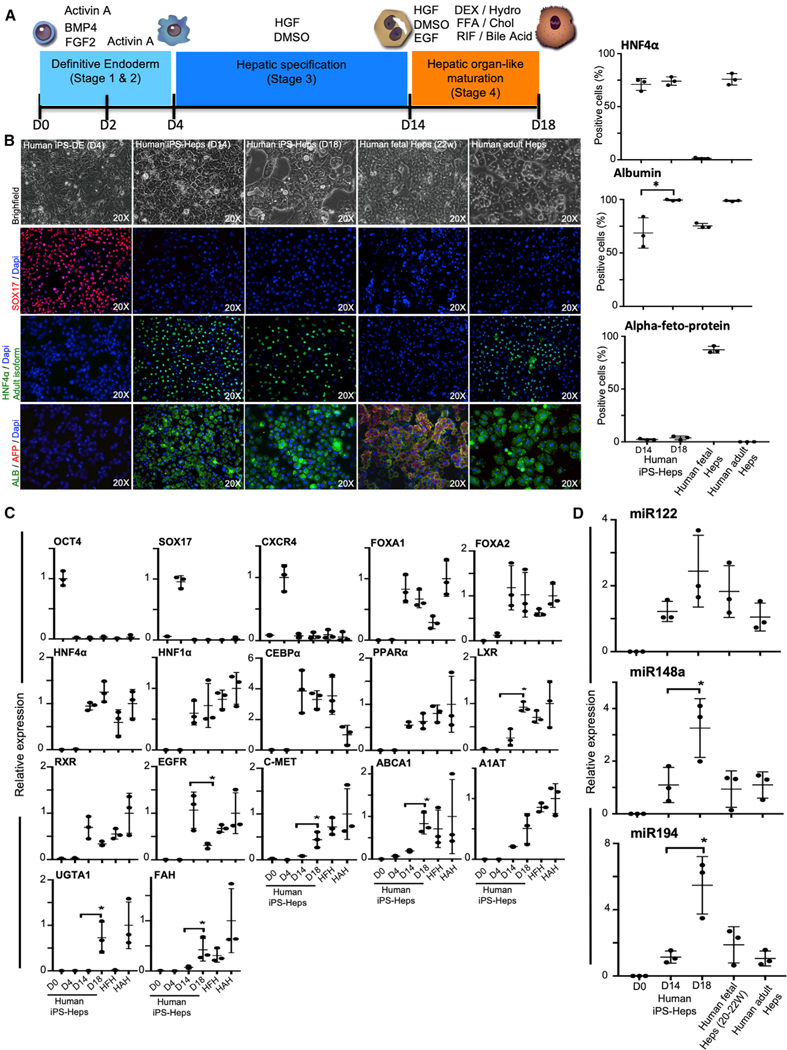
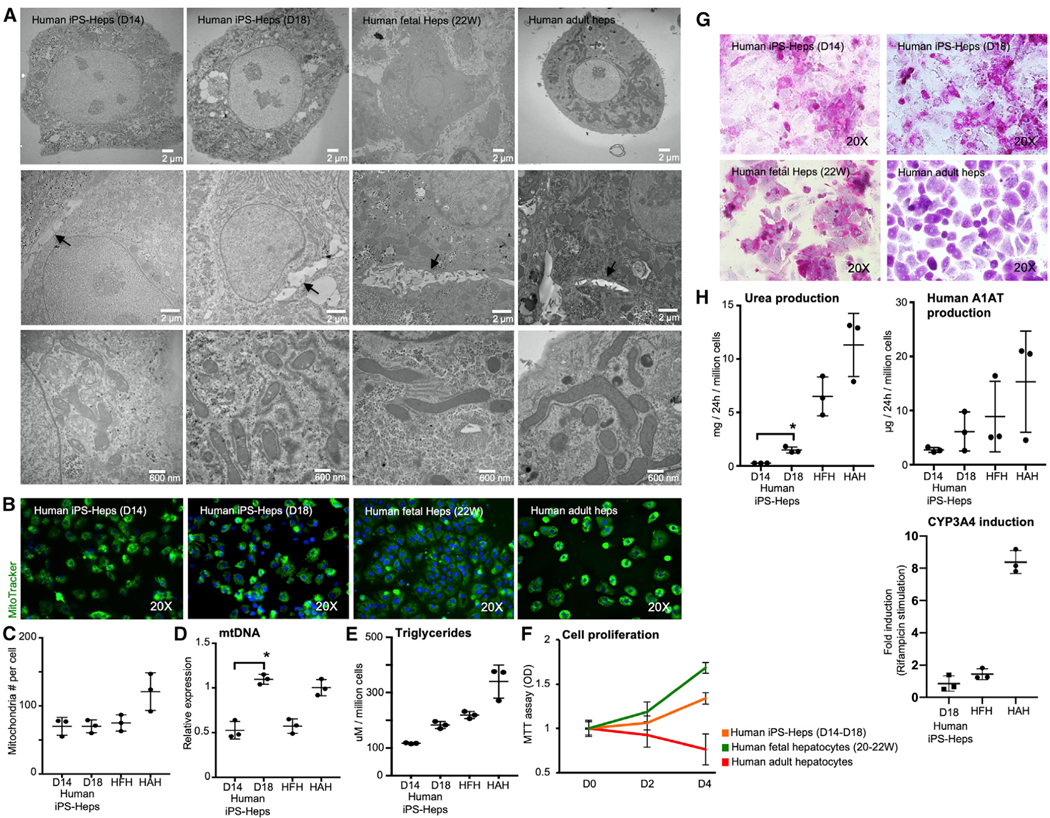
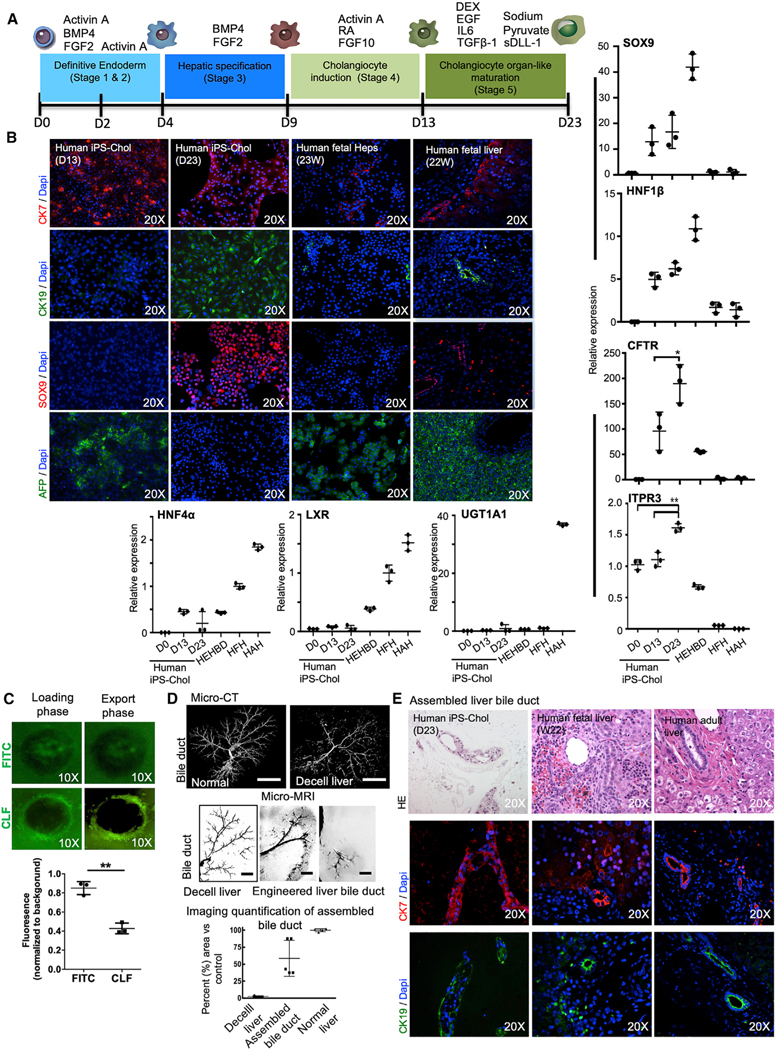
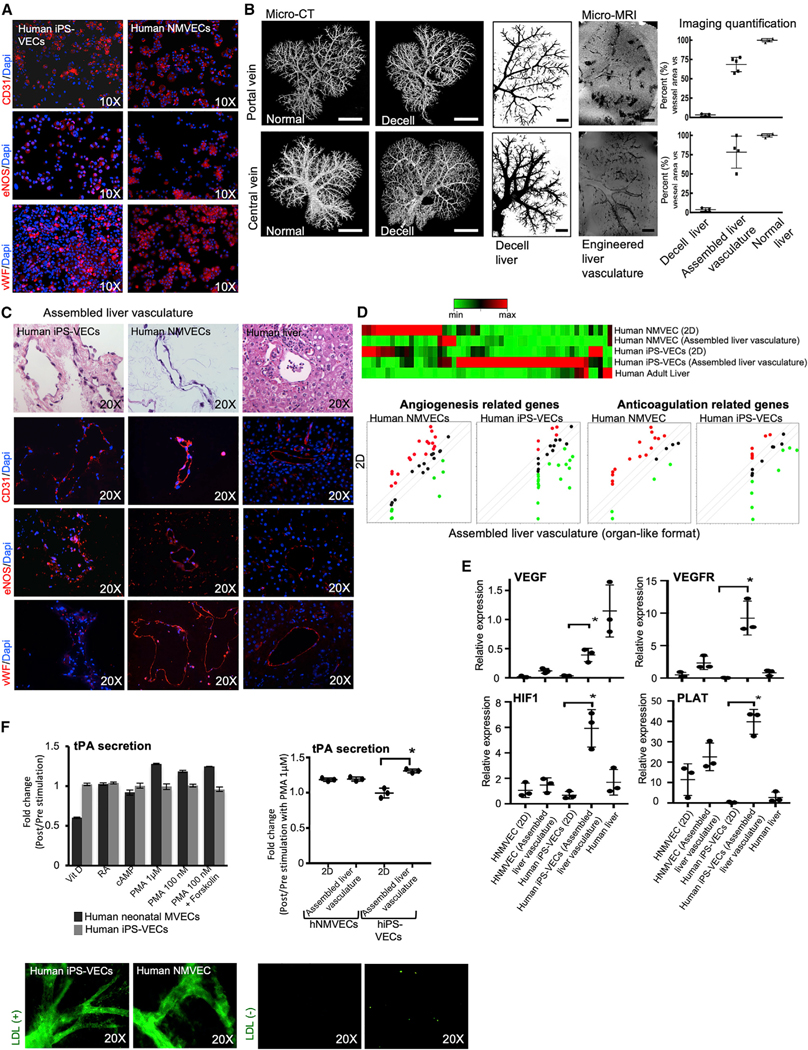
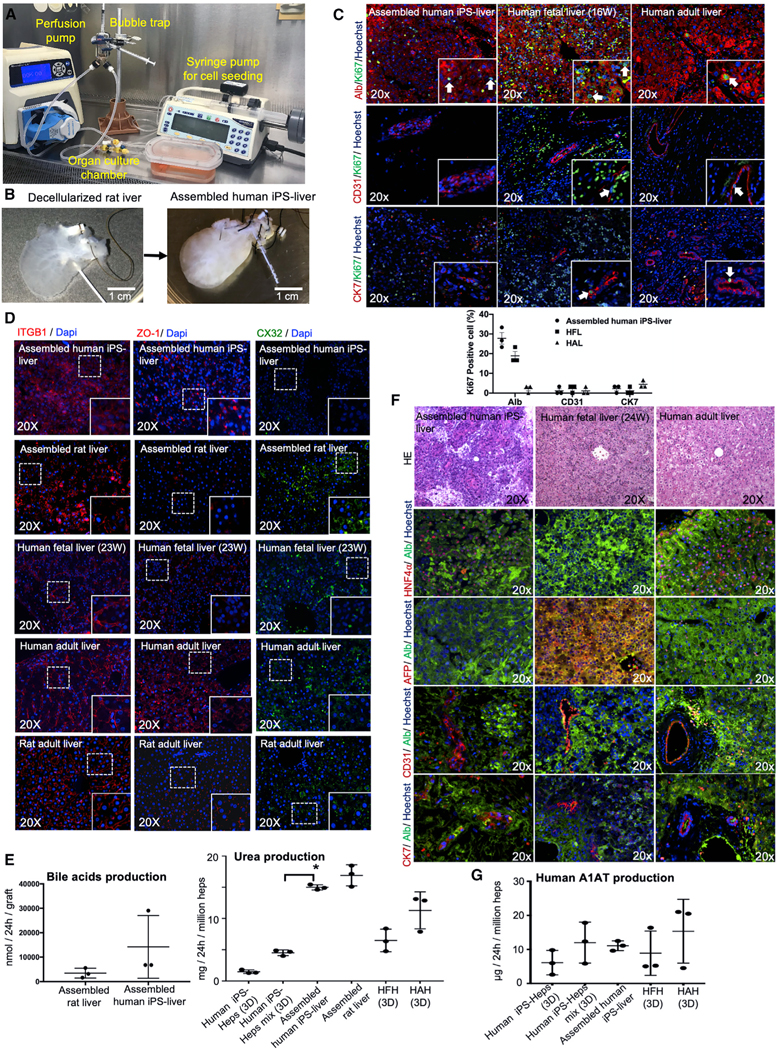
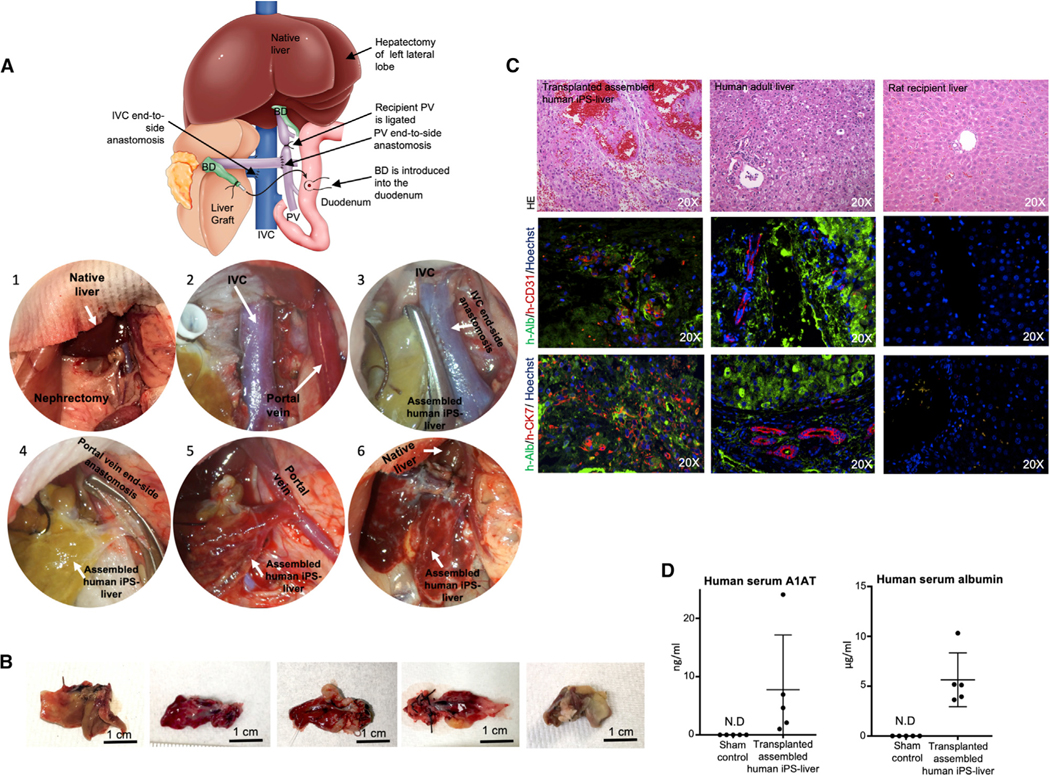
The availability of an autologous transplantable auxiliary liver would dramatically affect the treatment of liver disease. Assembly and function in vivo of a bioengineered human liver derived from induced pluripotent stem cells (iPSCs) has not been previously described. By improving methods for liver decellularization, recellularization, and differentiation of different liver cellular lineages of human iPSCs in an organ-like environment, we generated functional engineered human mini livers and performed transplantation in a rat model. Whereas previous studies recellularized liver scaffolds largely with rodent hepatocytes, we repopulated not only the parenchyma with human iPSC-hepatocytes but also the vascular system with human iPS-endothelial cells, and the bile duct network with human iPSC-biliary epithelial cells. The regenerated human iPSC-derived mini liver containing multiple cell types was tested in vivo and remained functional for 4 days after auxiliary liver transplantation in immunocompromised, engineered (IL2rg-/-) rats.
Keywords: bioengineered human liver; human iPS cells; human iPS-biliary cells; human iPS-endothelial cells; human iPS-hepatocytes; liver maturation; mini human liver; organ-microenvironment; transplantation.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests H.Y. and A.S.-Gare inventors on a patent application that involves some of the perfusion technology used in this work (WO/2011/002926); K.H., K.M., J.G.-L., H.Y., and A.S.-G. have an international patent related to this work that describes methods of preparing artificial organs and related compositions for transplantation and regeneration (WO/2015/168254). K.T., A.C.H., J.G.-L., Y.W., T.M., and A.S.-G. have a provisional international patent application that describes hepatic differentiation of human pluripotent stem cells and liver repopulation (PCT/US2018/018032). A.S.-G., J.G.-L., K.T., A.C.H., Y.W., T.M., and I.J.F. are co-founders and have a financial interest in Von Baer Wolff, Inc. a company focused on biofabrication of autologous human hepatocytes from stem cells technology and programming liver failure and their interests are managed by the Conflict of Interest Office at the University of Pittsburgh in accordance with their policies.
Figures
Comment in
-
Bioengineered Mini Human Livers for Transplantation.Hepatology. 2021 Jan;73(1):449-451. doi: 10.1002/hep.31528. Epub 2020 Dec 10. Hepatology. 2021. PMID: 32865234 No abstract available.
References
-
- Ammori JB, Pelletier SJ, Lynch R, Cohn J, Ads Y, Campbell DA, and Englesbe MJ (2008). Incremental costs of post-liver transplantation complications. J. Am. Coll. Surg 206, 89–95. - PubMed
-
- Avior Y, Levy G, Zimerman M, Kitsberg D, Schwartz R, Sadeh R, Moussaieff A, Cohen M, Itskovitz-Eldor J, and Nahmias Y. (2015). Microbial-derived lithocholic acid and vitamin K2 drive the metabolic maturation of pluripotent stem cells-derived and fetal hepatocytes. Hepatology 62, 265–278. - PubMed
-
- Bandiera S, Pfeffer S, Baumert TF, and Zeisel MB (2015). miR-122–a key factor and therapeutic target in liver disease. J. Hepatol 62, 448–457. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
