

An Updated Overview on the Regulation of Seed Germination
- PMID: 32492790
- PMCID: PMC7356954
- DOI: 10.3390/plants9060703
An Updated Overview on the Regulation of Seed Germination
Abstract
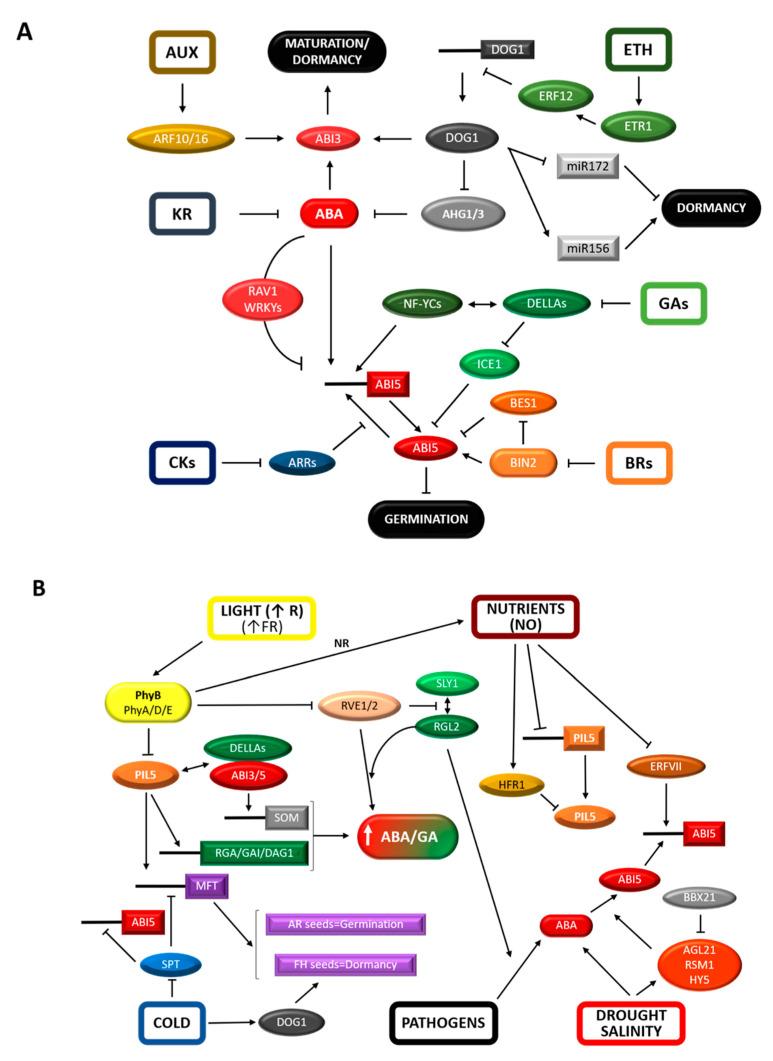
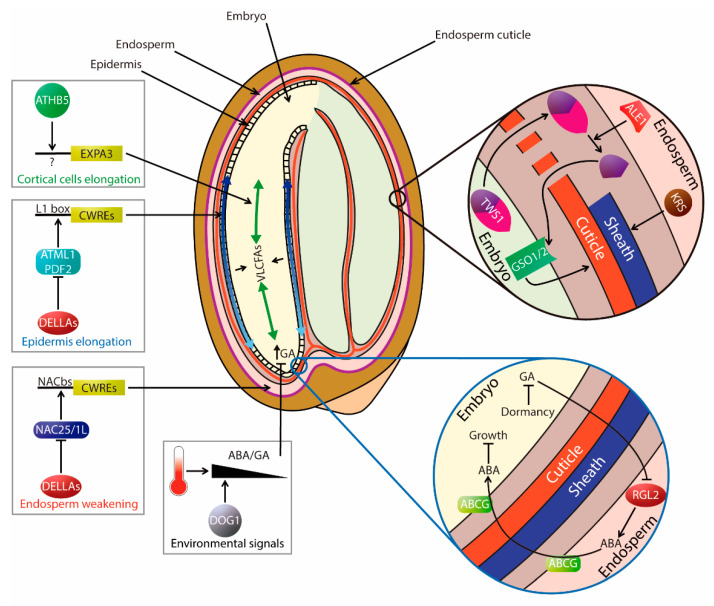
The ability of a seed to germinate and establish a plant at the right time of year is of vital importance from an ecological and economical point of view. Due to the fragility of these early growth stages, their swiftness and robustness will impact later developmental stages and crop yield. These traits are modulated by a continuous interaction between the genetic makeup of the plant and the environment from seed production to germination stages. In this review, we have summarized the established knowledge on the control of seed germination from a molecular and a genetic perspective. This serves as a "backbone" to integrate the latest developments in the field. These include the link of germination to events occurring in the mother plant influenced by the environment, the impact of changes in the chromatin landscape, the discovery of new players and new insights related to well-known master regulators. Finally, results from recent studies on hormone transport, signaling, and biophysical and mechanical tissue properties are underscoring the relevance of tissue-specific regulation and the interplay of signals in this crucial developmental process.
Keywords: ABA/GA; environmental signals; epigenetics; hormone signaling and dynamics; post-transcriptional regulation; seed dormancy and germination; spatio-temporal regulation.; transcription factors.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Bewley J.D., Bradford K., Hilhorst H., Nonogaki H. Seeds, Physiology of Development, Germination and Dormancy. 3rd ed. Springer; New York, NY, USA: 2013. Dormancy and the control of germination; pp. 247–297.
-
- Willis C.G., Baskin C., Baskin J.M., Auld J.R., Venable D.L., Cavender-Bares J., Donohue K., De Casas R.R. The nescent germination working group the evolution of seed dormancy: Environmental cues, evolutionary hubs, and diversification of the seed plants. New Phytol. 2014;203:300–309. doi: 10.1111/nph.12782. - DOI - PubMed
-
- Koorneef M., Elgersma A., Hanhart C.J., Rijn L., Zeevaart J.A.D., Loenen-Martinet E.P. A gibberellin insensitive mutant of Arabidopsis thaliana. Physiol. Plant. 1985;65:33–39. doi: 10.1111/j.1399-3054.1985.tb02355.x. - DOI
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
