

Convergent genes shape budding yeast pericentromeres
- PMID: 32494069
- PMCID: PMC7279958
- DOI: 10.1038/s41586-020-2244-6
Convergent genes shape budding yeast pericentromeres
Abstract
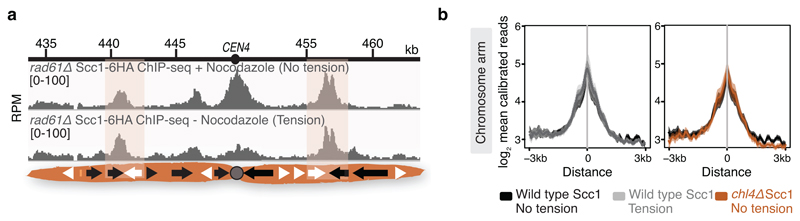
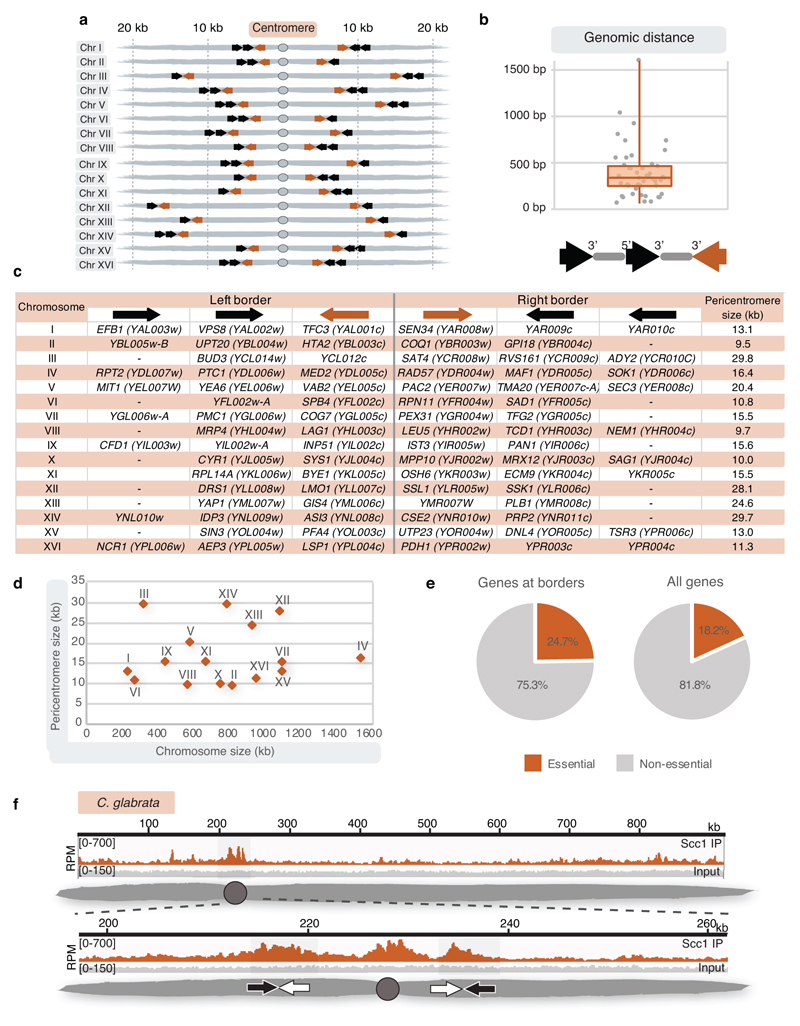
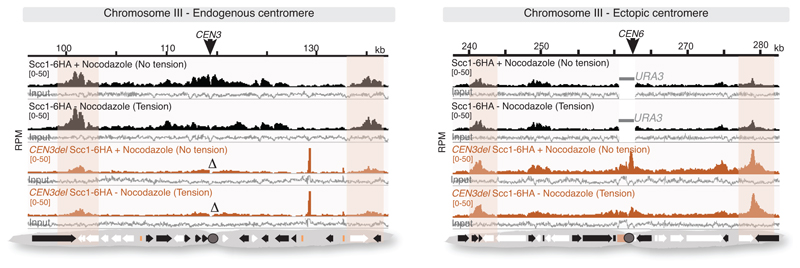
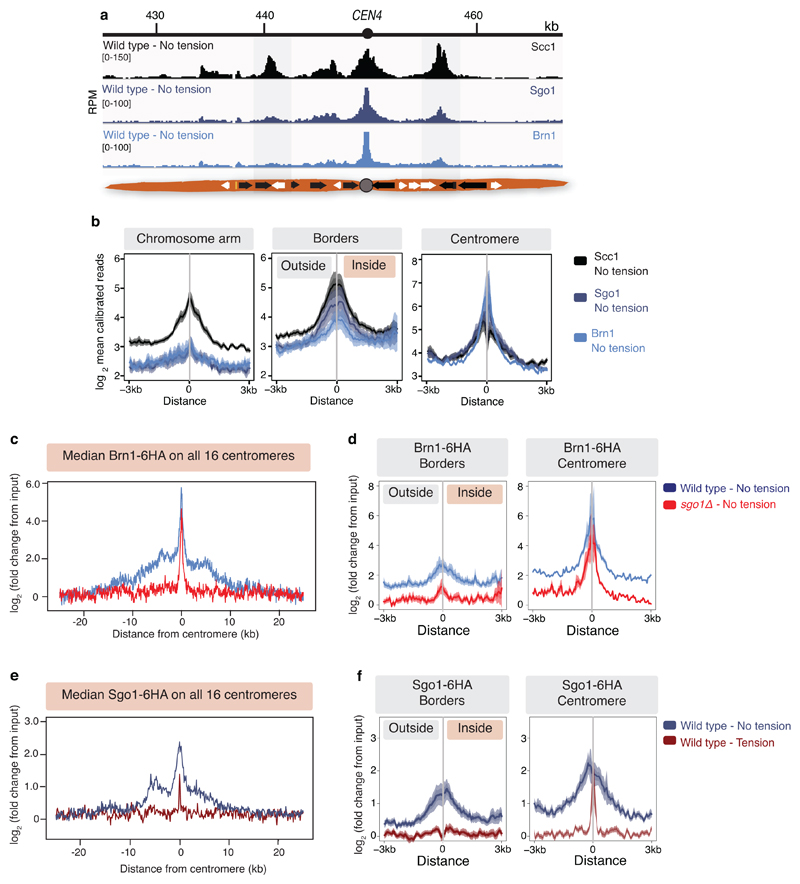
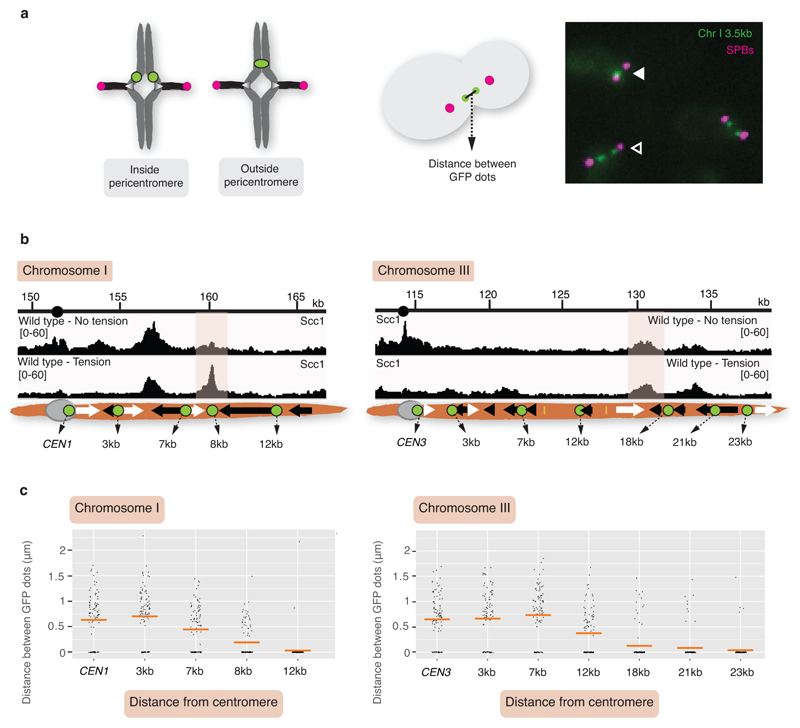
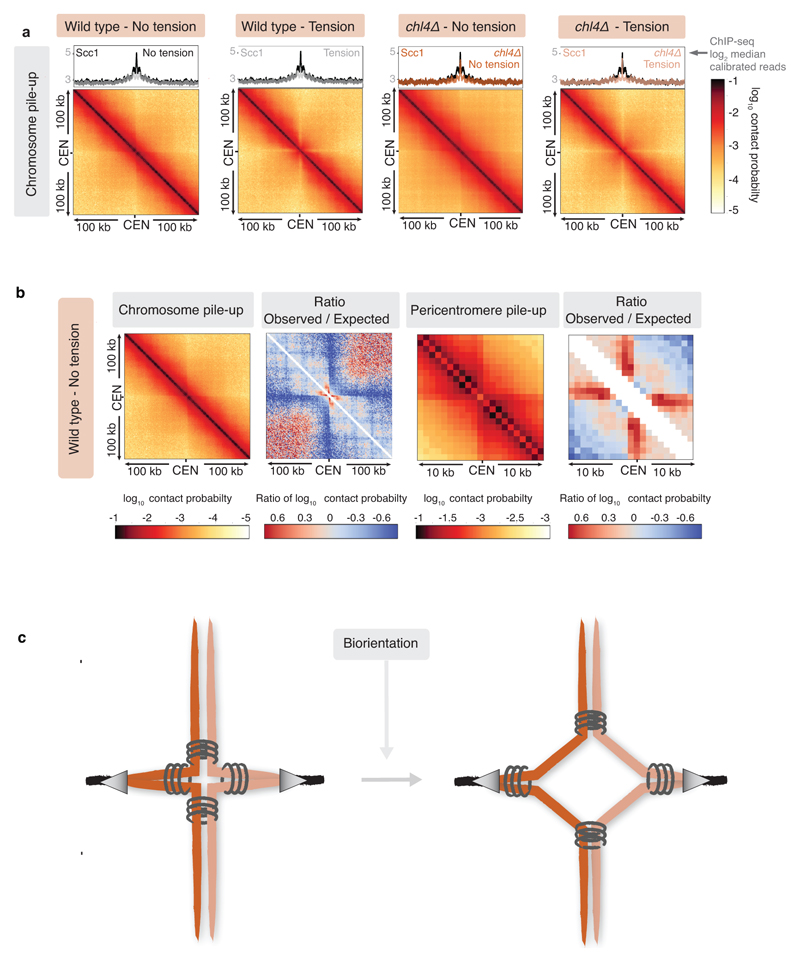
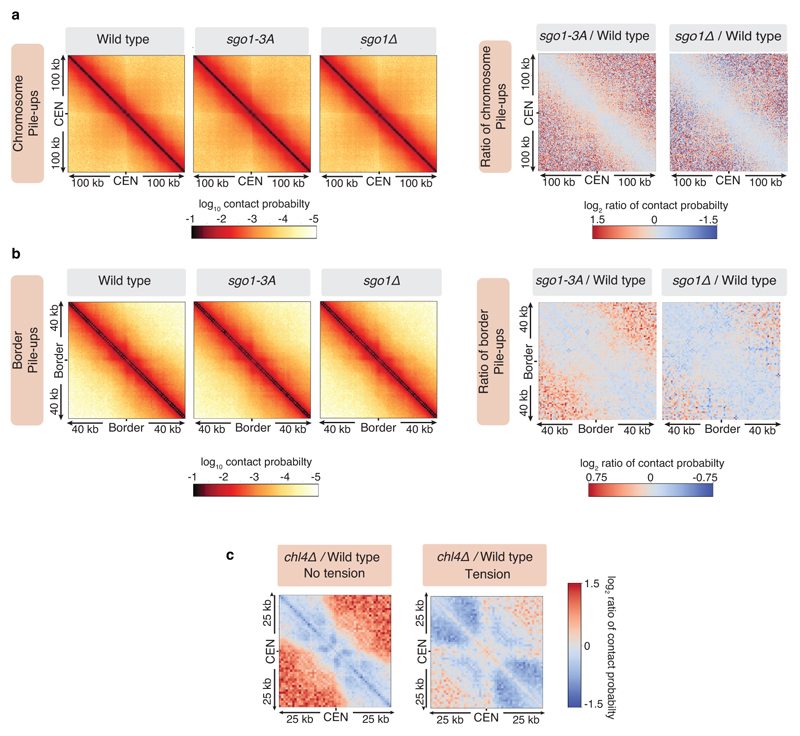
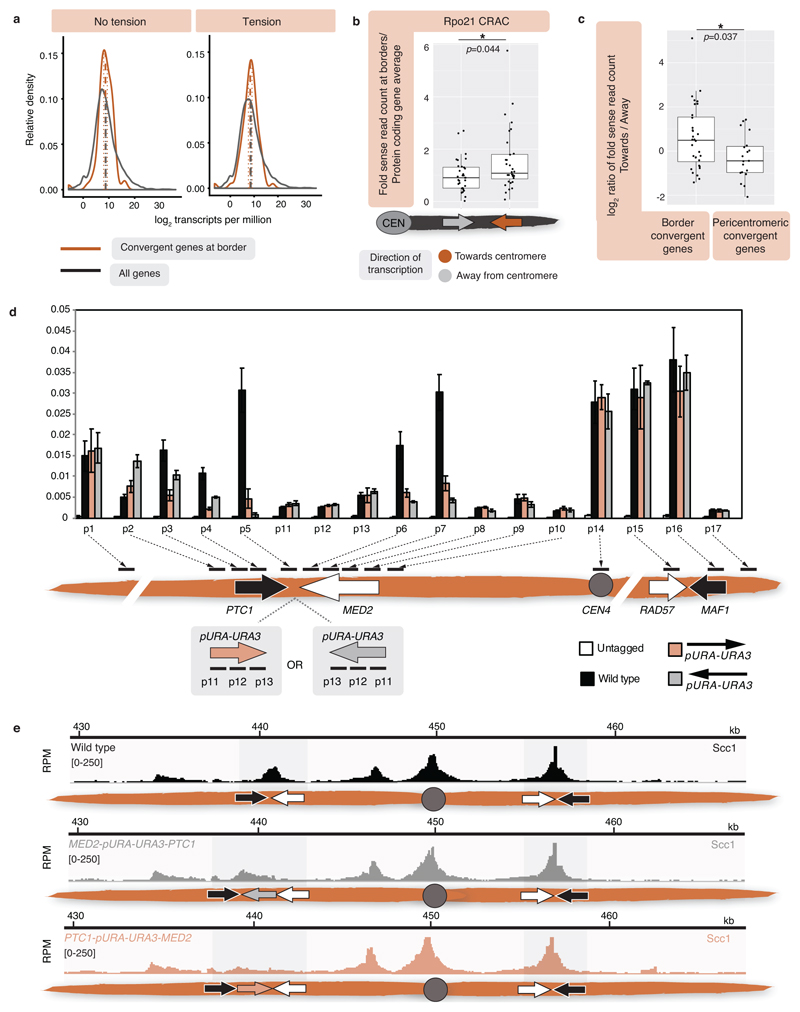
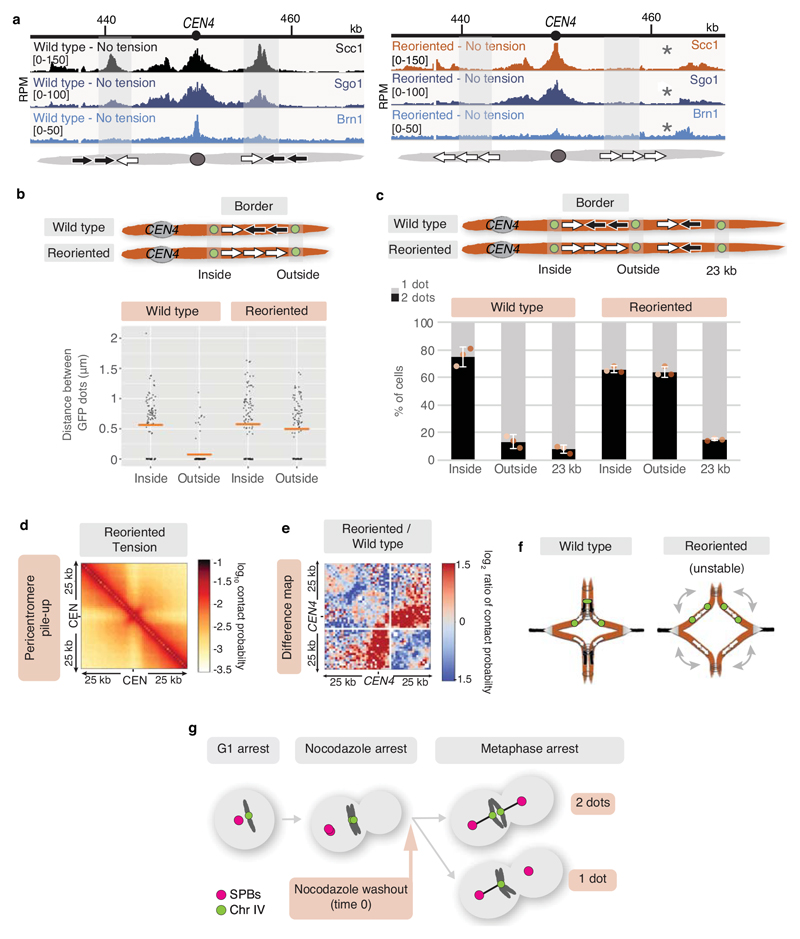
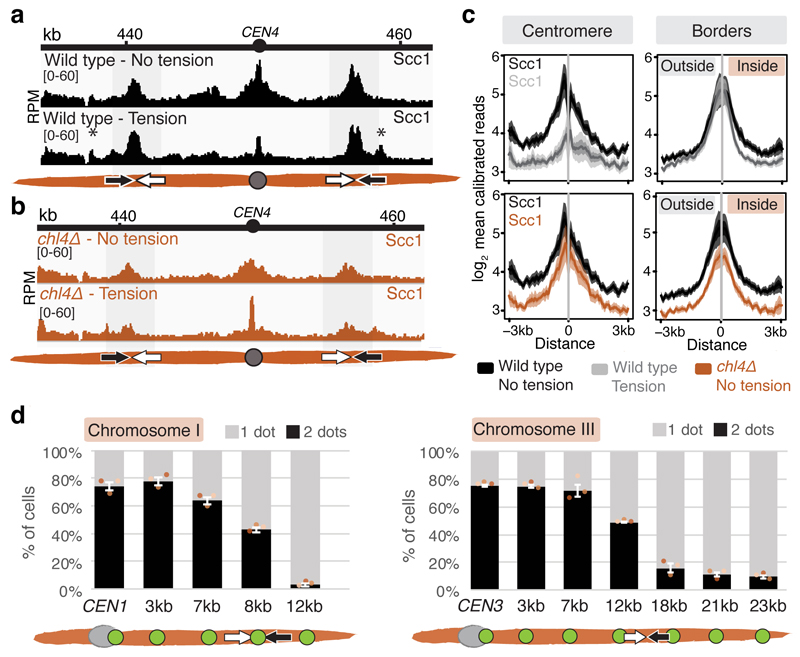
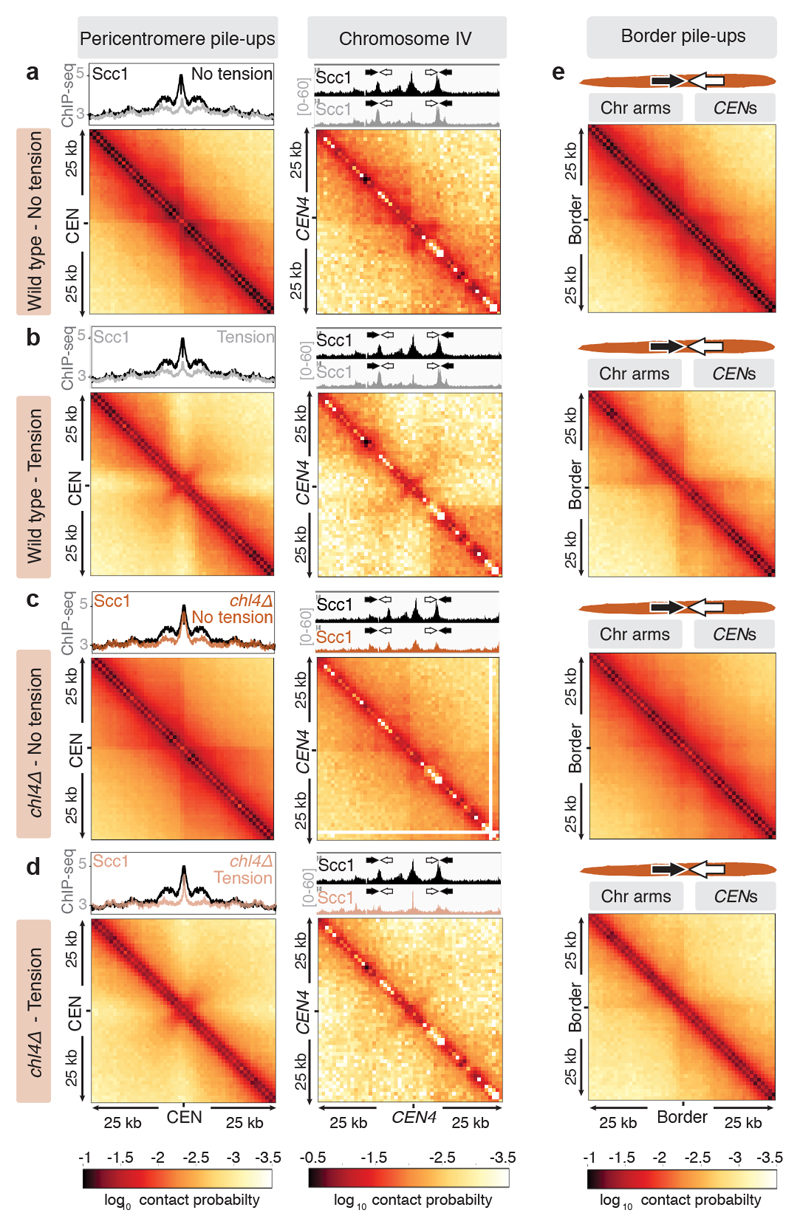
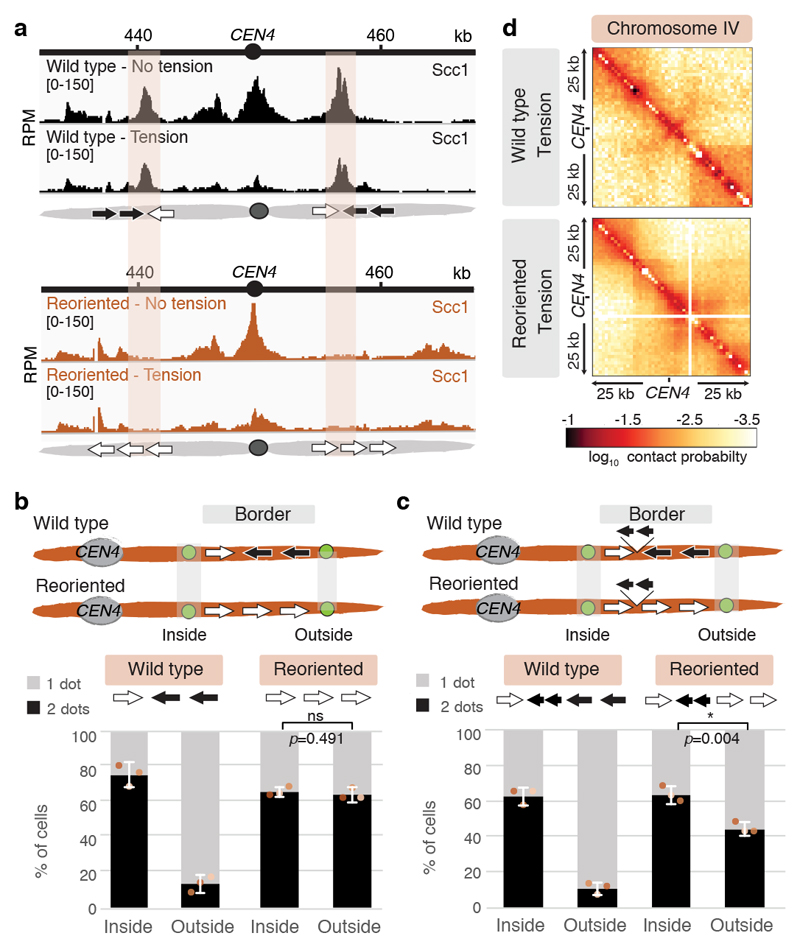
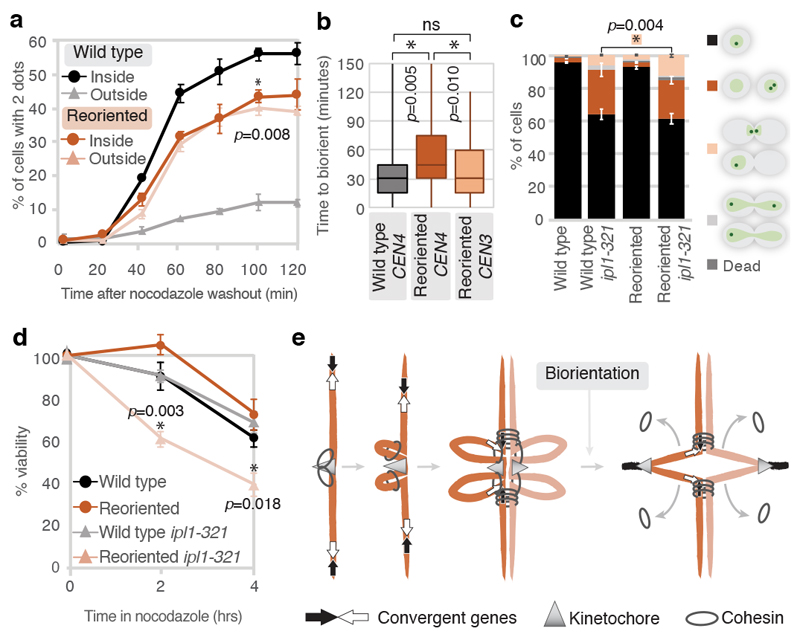
The three-dimensional architecture of the genome governs its maintenance, expression and transmission. The cohesin protein complex organizes the genome by topologically linking distant loci, and is highly enriched in specialized chromosomal domains surrounding centromeres, called pericentromeres1-6. Here we report the three-dimensional structure of pericentromeres in budding yeast (Saccharomyces cerevisiae) and establish the relationship between genome organization and function. We find that convergent genes mark pericentromere borders and, together with core centromeres, define their structure and function by positioning cohesin. Centromeres load cohesin, and convergent genes at pericentromere borders trap it. Each side of the pericentromere is organized into a looped conformation, with border convergent genes at the base. Microtubule attachment extends a single pericentromere loop, size-limited by convergent genes at its borders. Reorienting genes at borders into a tandem configuration repositions cohesin, enlarges the pericentromere and impairs chromosome biorientation during mitosis. Thus, the linear arrangement of transcriptional units together with targeted cohesin loading shapes pericentromeres into a structure that is competent for chromosome segregation. Our results reveal the architecture of the chromosomal region within which kinetochores are embedded, as well as the restructuring caused by microtubule attachment. Furthermore, we establish a direct, causal relationship between the three-dimensional genome organization of a specific chromosomal domain and cellular function.
Conflict of interest statement
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
