

Structure and mechanism of the mitochondrial Ca2+ uniporter holocomplex
- PMID: 32494073
- PMCID: PMC7544431
- DOI: 10.1038/s41586-020-2309-6
Structure and mechanism of the mitochondrial Ca2+ uniporter holocomplex
Abstract
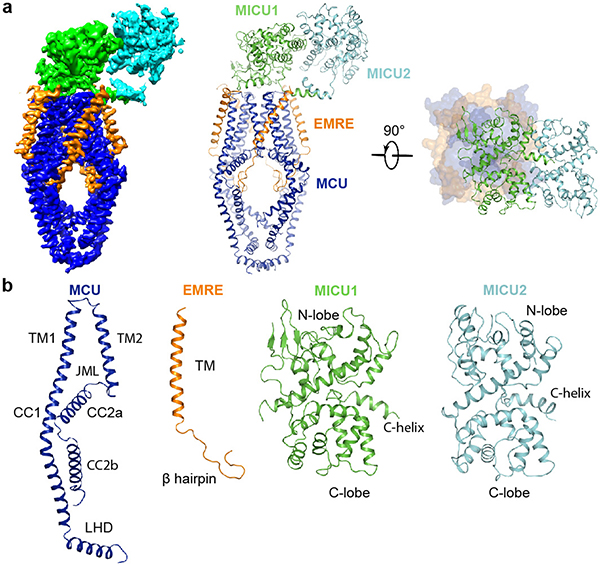
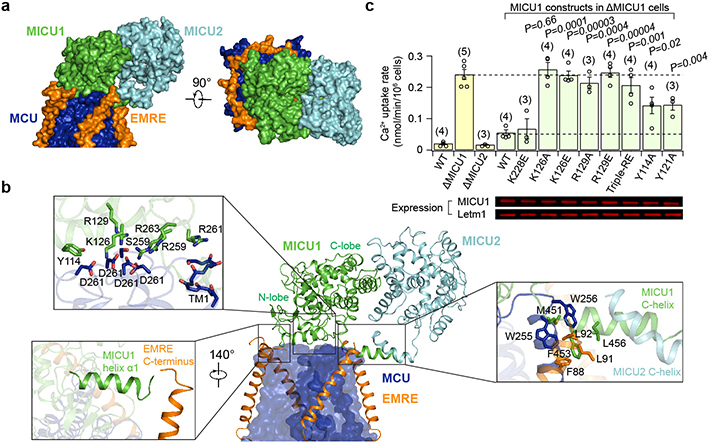
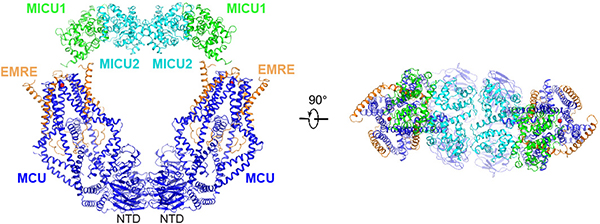
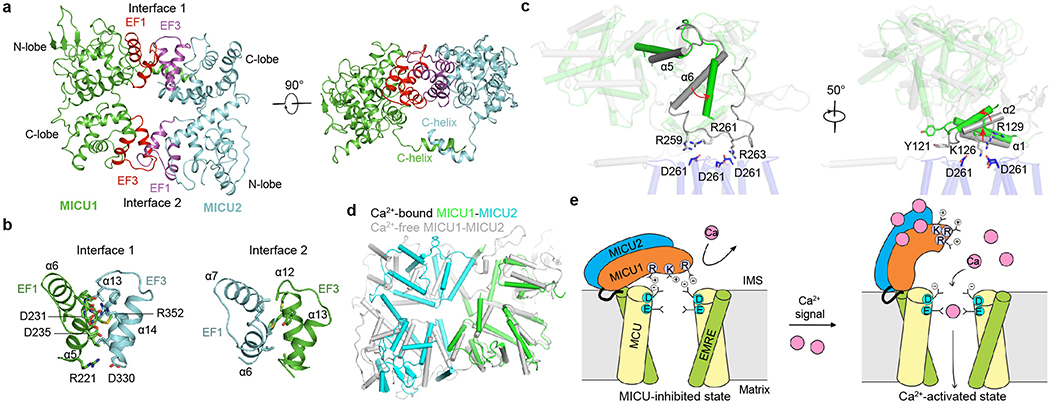
Mitochondria take up Ca2+ through the mitochondrial calcium uniporter complex to regulate energy production, cytosolic Ca2+ signalling and cell death1,2. In mammals, the uniporter complex (uniplex) contains four core components: the pore-forming MCU protein, the gatekeepers MICU1 and MICU2, and an auxiliary subunit, EMRE, essential for Ca2+ transport3-8. To prevent detrimental Ca2+ overload, the activity of MCU must be tightly regulated by MICUs, which sense changes in cytosolic Ca2+ concentrations to switch MCU on and off9,10. Here we report cryo-electron microscopic structures of the human mitochondrial calcium uniporter holocomplex in inhibited and Ca2+-activated states. These structures define the architecture of this multicomponent Ca2+-uptake machinery and reveal the gating mechanism by which MICUs control uniporter activity. Our work provides a framework for understanding regulated Ca2+ uptake in mitochondria, and could suggest ways of modulating uniporter activity to treat diseases related to mitochondrial Ca2+ overload.
Conflict of interest statement
Figures
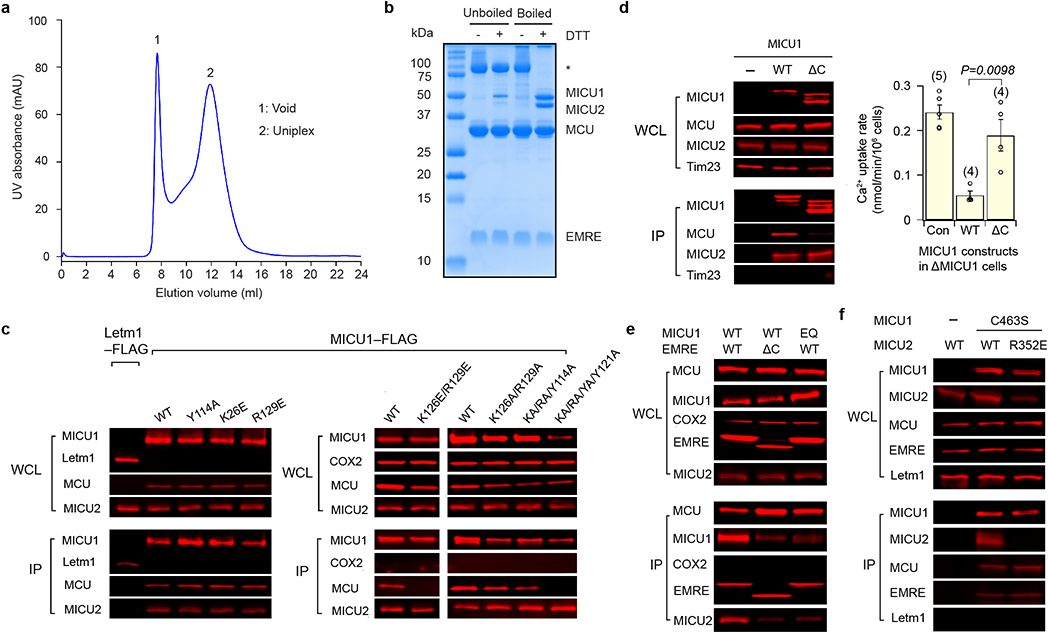
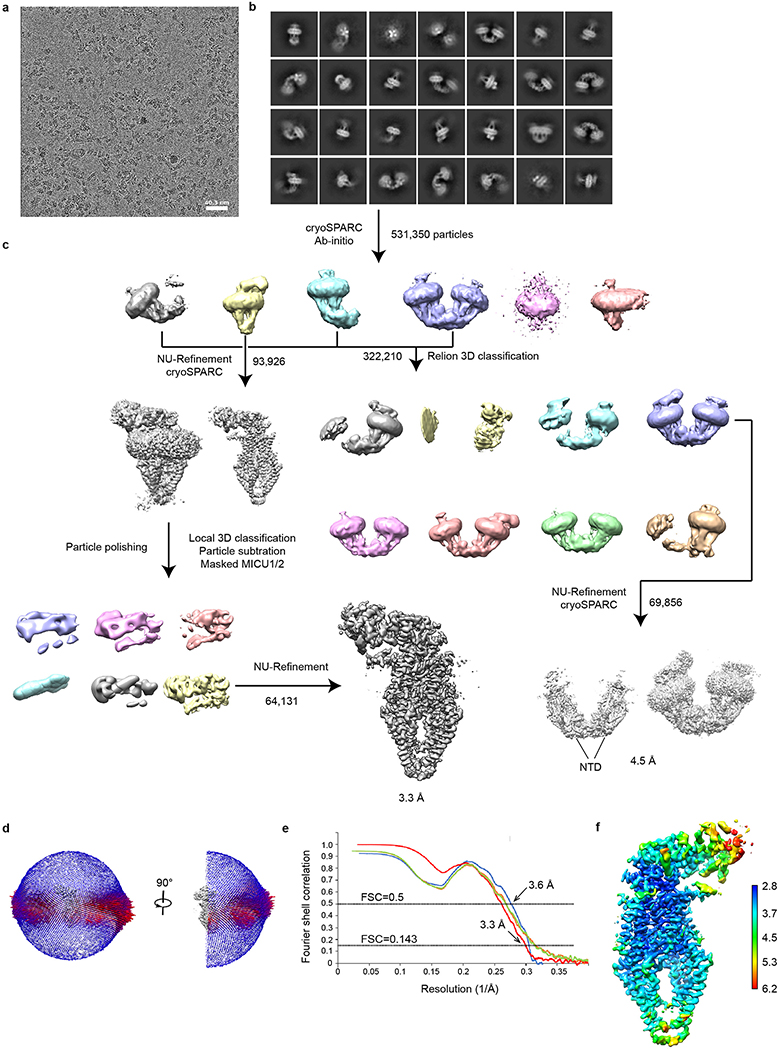

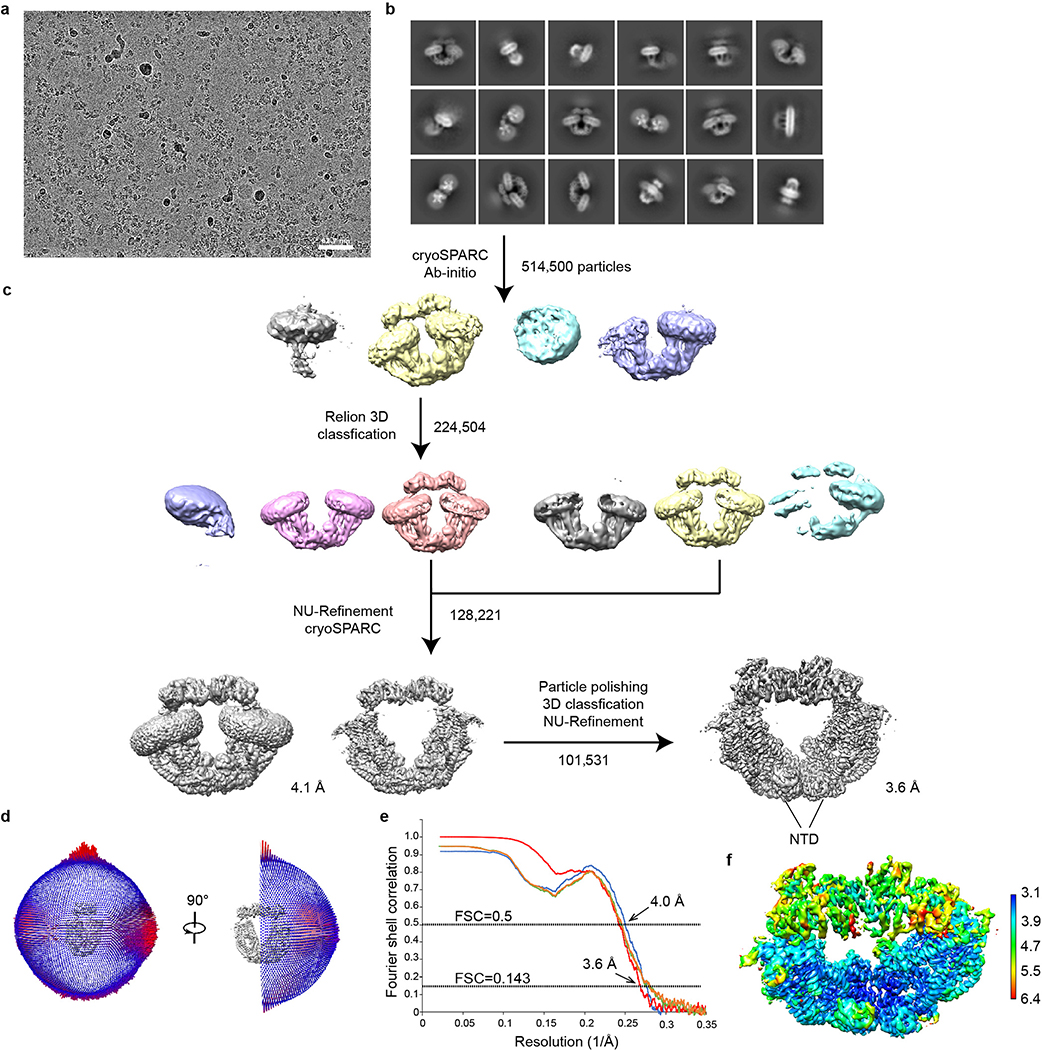
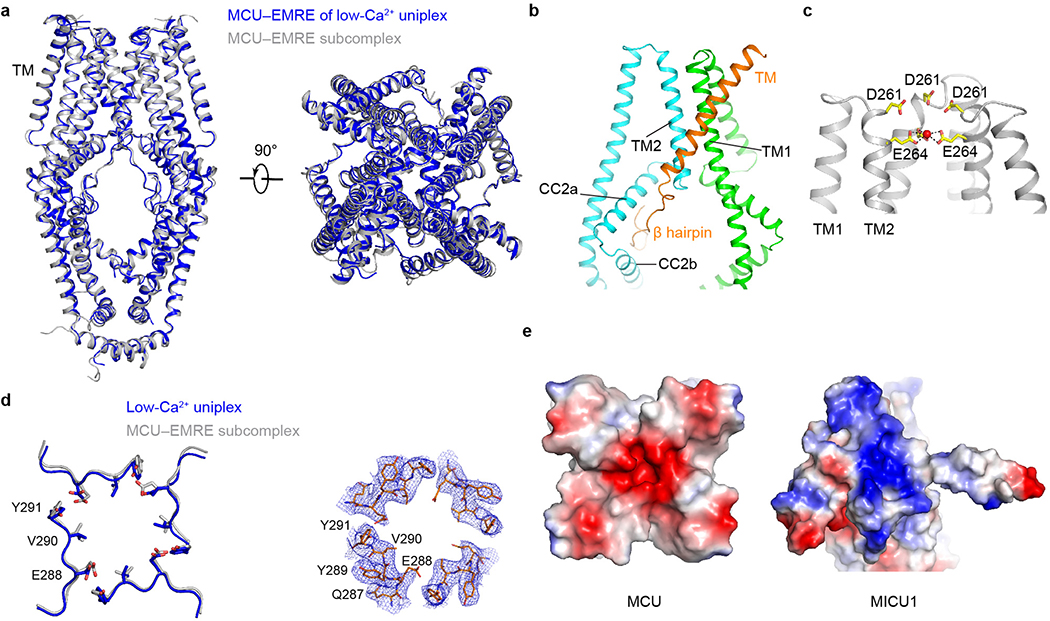
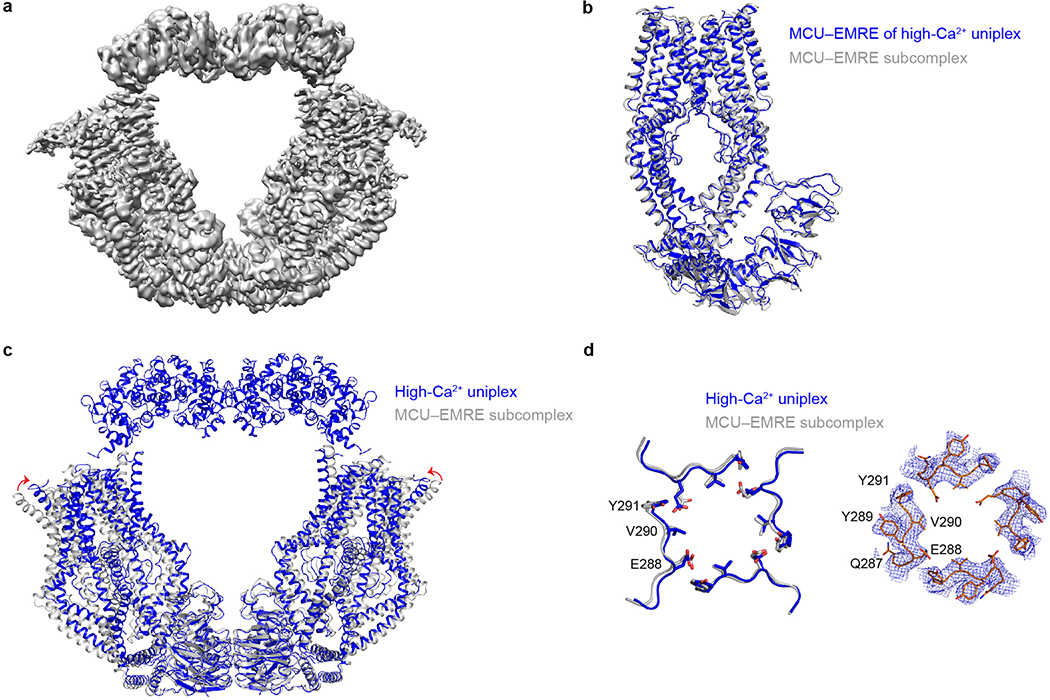
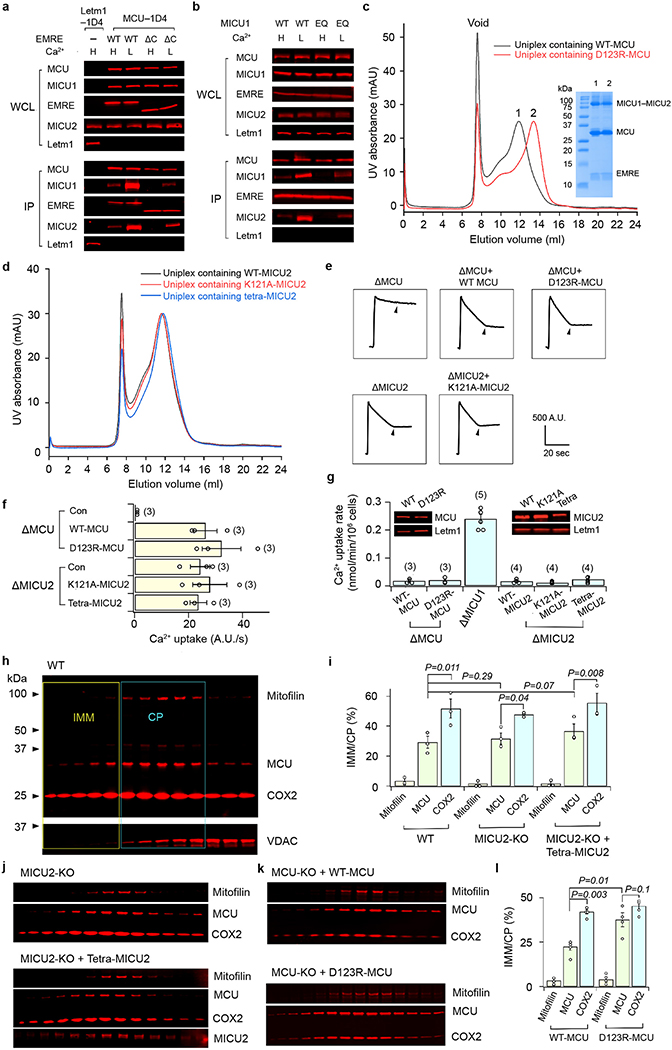
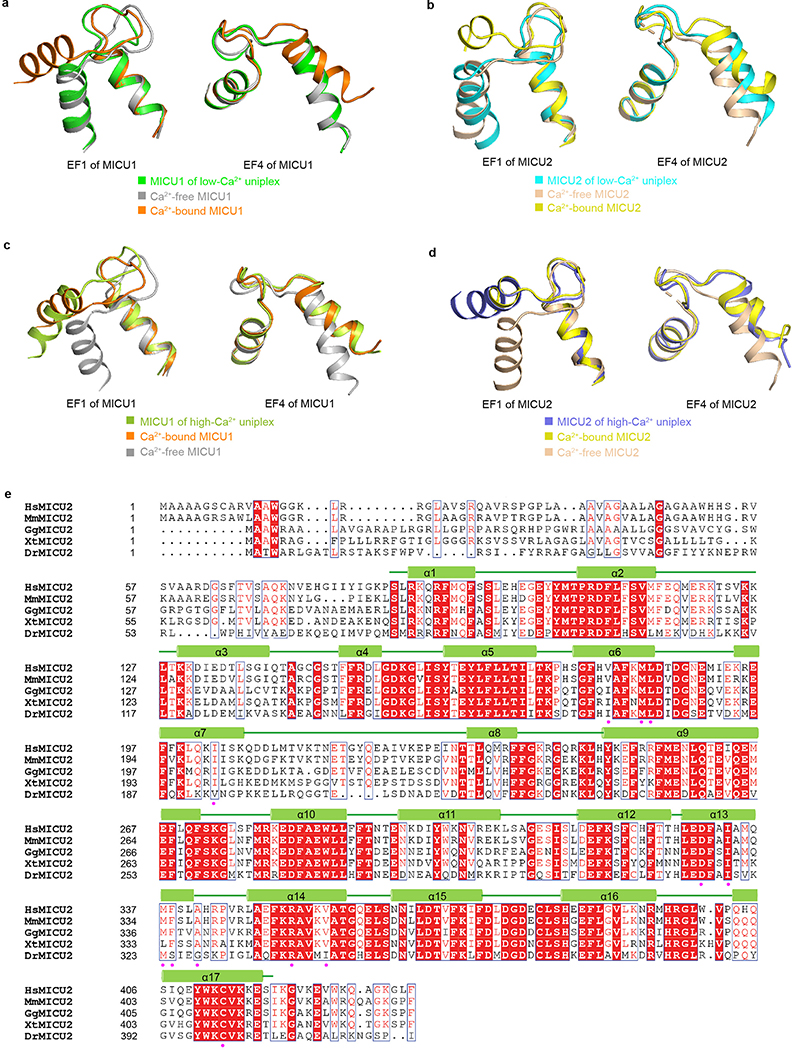
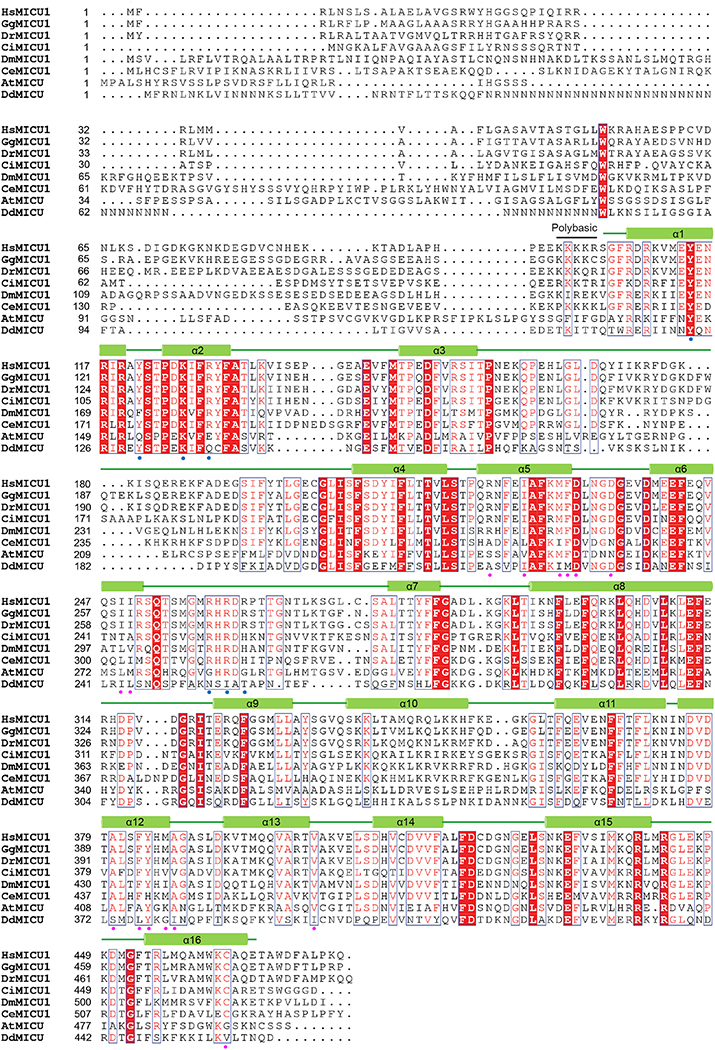
References
-
- Rizzuto R, De Stefani D, Raffaello A & Mammucari C Mitochondria as sensors and regulators of calcium signalling. Nat. Rev. Mol. Cell Biol 13, 566–578 (2012). - PubMed
-
- Giorgi C, Marchi S & Pinton P The machineries, regulation and cellular functions of mitochondrial calcium. Nat. Rev. Mol. Cell Biol 19, 713–730 (2018). - PubMed
-
- Kirichok Y, Krapivinsky G & Clapham DE The mitochondrial calcium uniporter is a highly selective ion channel. Nature 427, 360–364 (2004). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
