

Nothing Is Yet Set in (Hi)stone: Novel Post-Translational Modifications Regulating Chromatin Function
- PMID: 32498971
- PMCID: PMC7502514
- DOI: 10.1016/j.tibs.2020.05.009
Nothing Is Yet Set in (Hi)stone: Novel Post-Translational Modifications Regulating Chromatin Function
Abstract
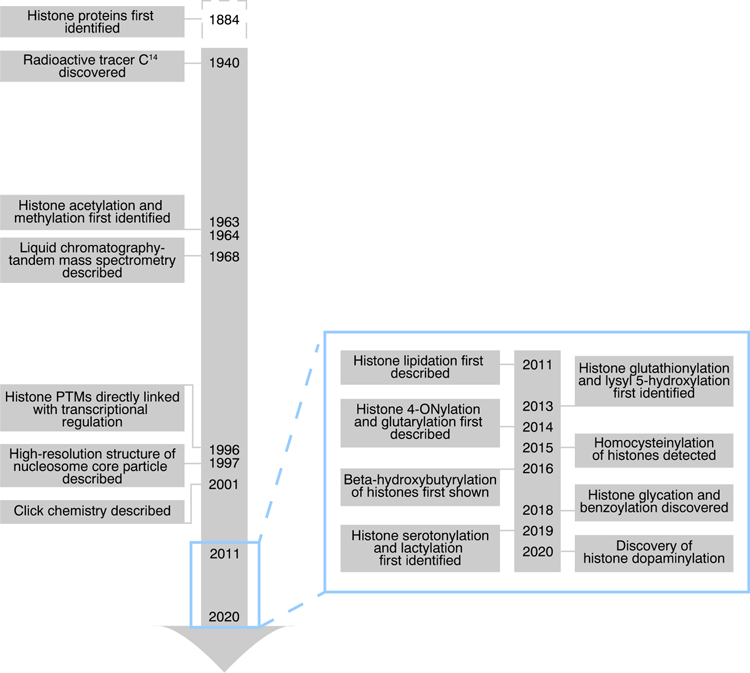
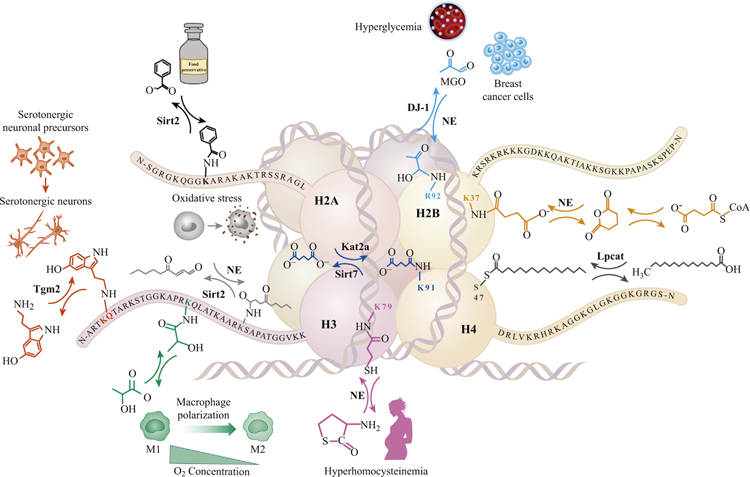
Histone post-translational modifications (PTMs) have emerged as exciting mechanisms of biological regulation, impacting pathways related to cancer, immunity, brain function, and more. Over the past decade alone, several histone PTMs have been discovered, including acylation, lipidation, monoaminylation, and glycation, many of which appear to have crucial roles in nucleosome stability and transcriptional regulation. In this review, we discuss novel histone PTMs identified within the past 10 years, with an extended focus on enzymatic versus nonenzymatic mechanisms underlying modification and adduction. Furthermore, we consider how these novel histone PTMs might fit within the framework of a so-called 'histone code', emphasizing the physiological relevance of these PTMs in metabolism, development, and disease states.
Keywords: acylation; glycation; histone code hypothesis; lipidation; monoaminylation; nonenzymatic adduction.
Copyright © 2020 Elsevier Ltd. All rights reserved.
Figures
References
-
- Kornberg RD et al. (1999) Twenty-five years of the nucleosome, fundamental particle of the eukaryote chromosome. Cell 98, 285–294 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
