

Nutrient-dependent control of RNA polymerase II elongation rate regulates specific gene expression programs by alternative polyadenylation
- PMID: 32499400
- PMCID: PMC7328516
- DOI: 10.1101/gad.337212.120
Nutrient-dependent control of RNA polymerase II elongation rate regulates specific gene expression programs by alternative polyadenylation
Abstract
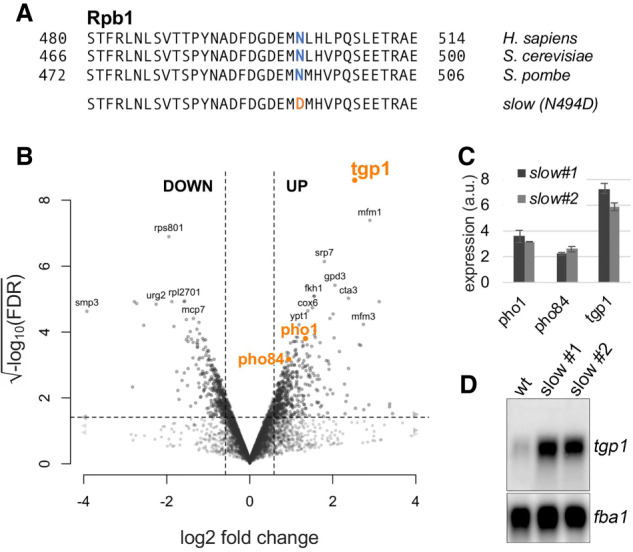
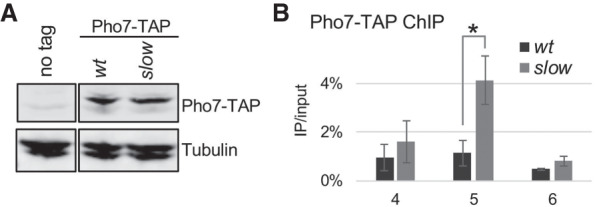
Transcription by RNA polymerase II (RNAPII) is a dynamic process with frequent variations in the elongation rate. However, the physiological relevance of variations in RNAPII elongation kinetics has remained unclear. Here we show in yeast that a RNAPII mutant that reduces the transcription elongation rate causes widespread changes in alternative polyadenylation (APA). We unveil two mechanisms by which APA affects gene expression in the slow mutant: 3' UTR shortening and gene derepression by premature transcription termination of upstream interfering noncoding RNAs. Strikingly, the genes affected by these mechanisms are enriched for functions involved in phosphate uptake and purine synthesis, processes essential for maintenance of the intracellular nucleotide pool. As nucleotide concentration regulates transcription elongation, our findings argue that RNAPII is a sensor of nucleotide availability and that genes important for nucleotide pool maintenance have adopted regulatory mechanisms responsive to reduced rates of transcription elongation.
Keywords: NTP sensing; RNA polymerase II; alternative polyadenylation; phosphate starvation; transcription elongation rate; transcription termination.
© 2020 Yague-Sanz et al.; Published by Cold Spring Harbor Laboratory Press.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials