

poly(UG)-tailed RNAs in genome protection and epigenetic inheritance
- PMID: 32499657
- PMCID: PMC8396162
- DOI: 10.1038/s41586-020-2323-8
poly(UG)-tailed RNAs in genome protection and epigenetic inheritance
Erratum in
-
Author Correction: poly(UG)-tailed RNAs in genome protection and epigenetic inheritance.Nature. 2021 Apr;592(7856):E27. doi: 10.1038/s41586-021-03364-y. Nature. 2021. PMID: 33854240 No abstract available.
Abstract
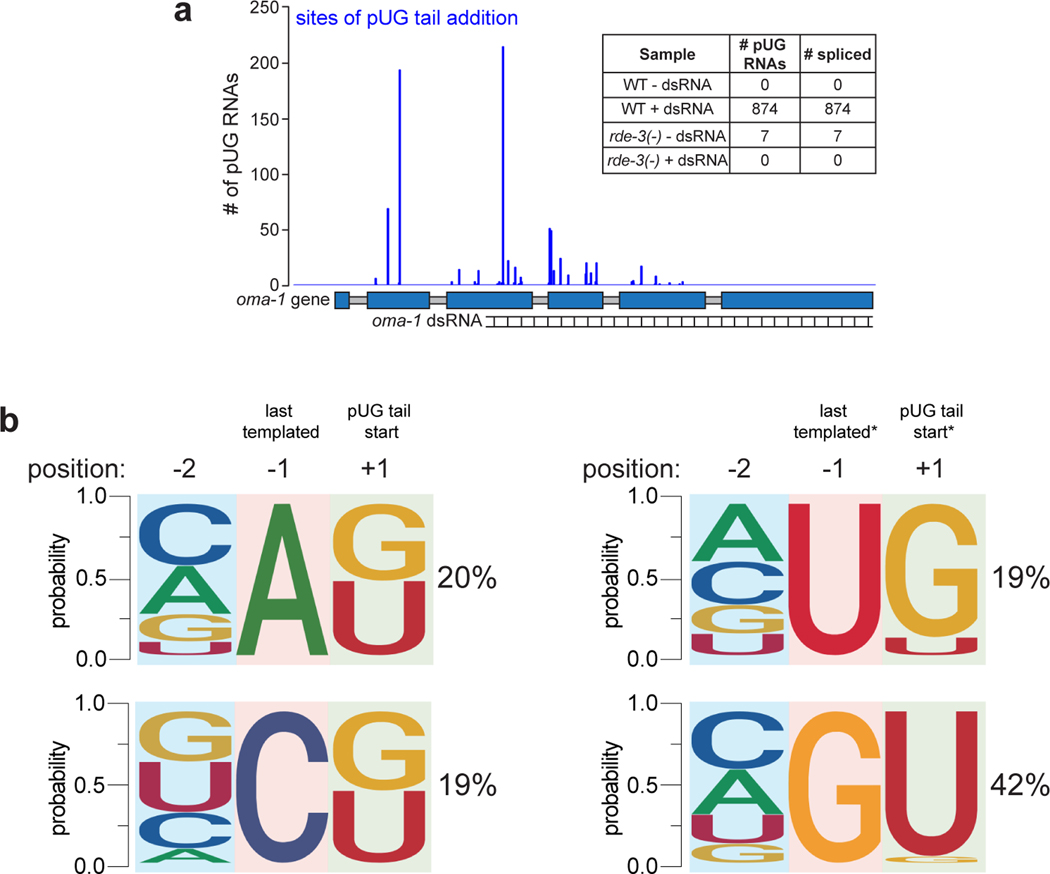
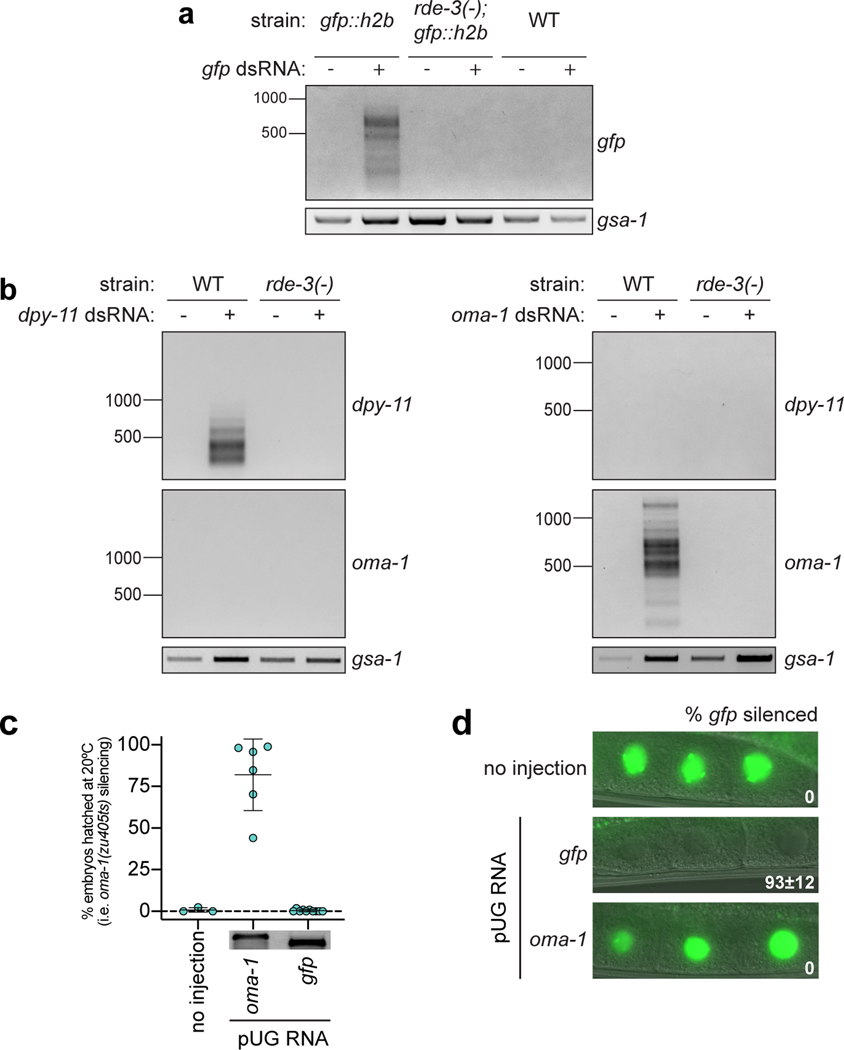
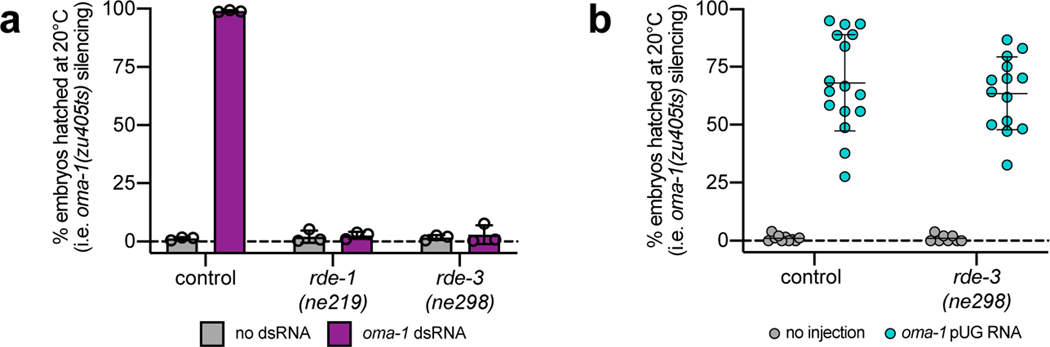
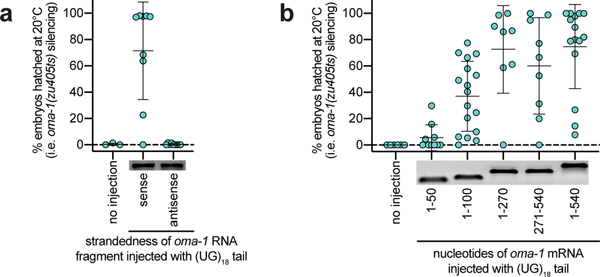
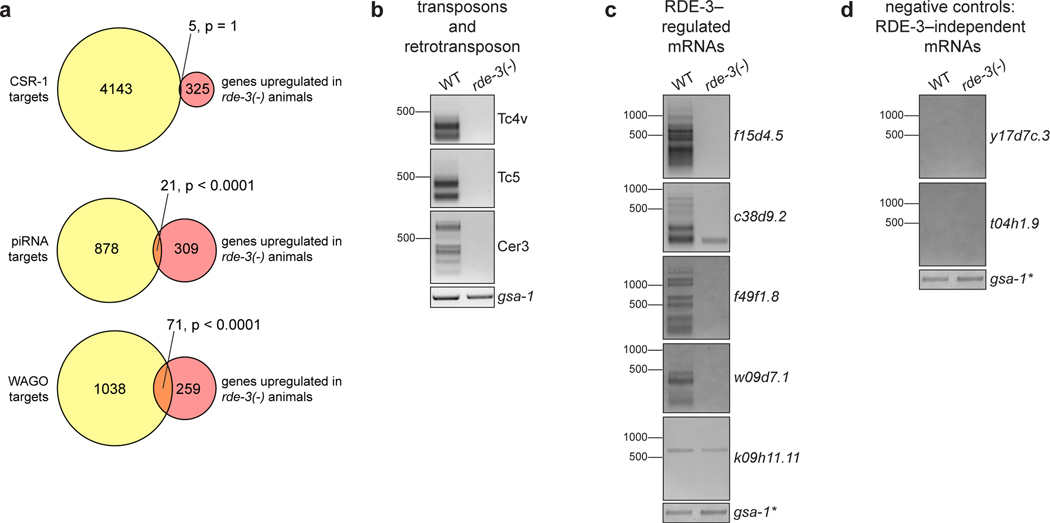
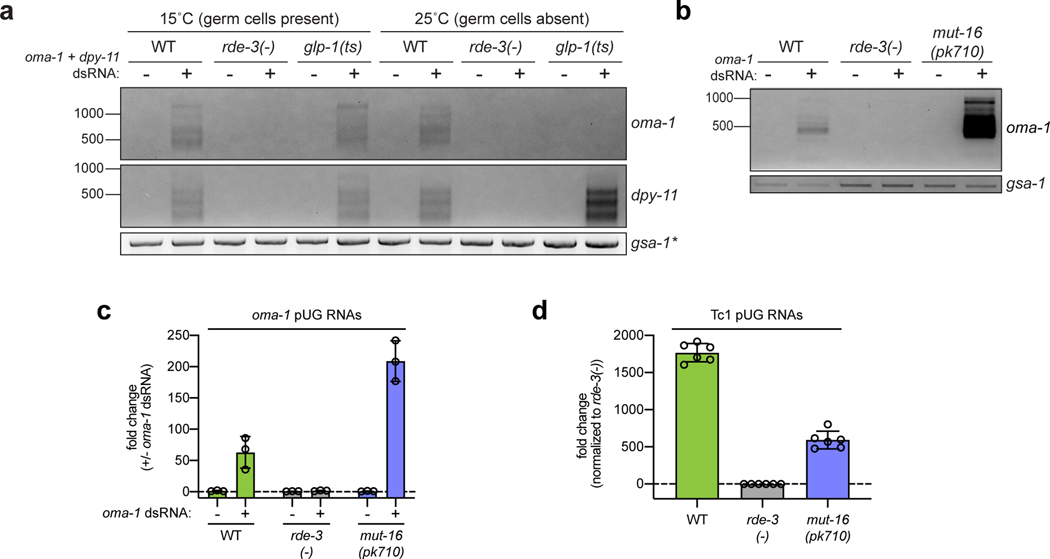
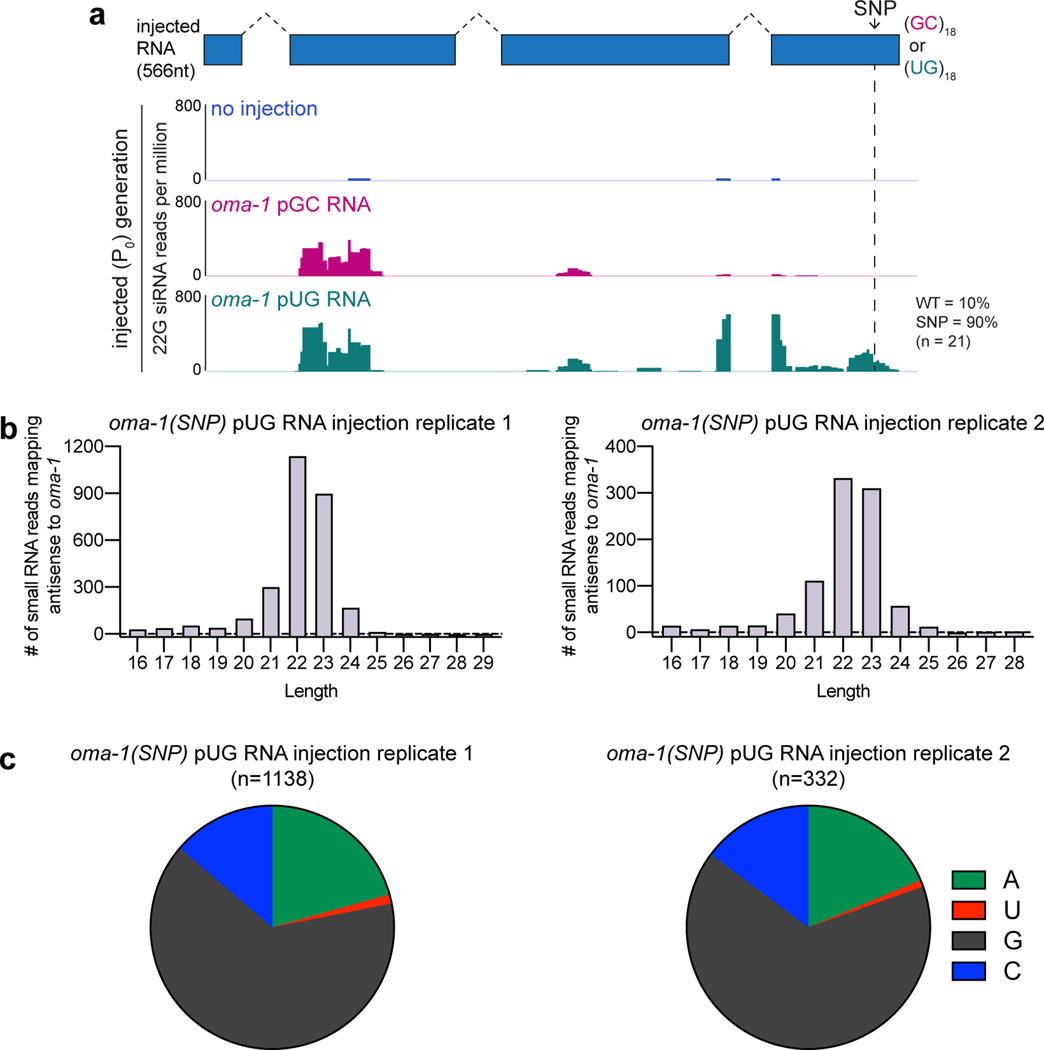
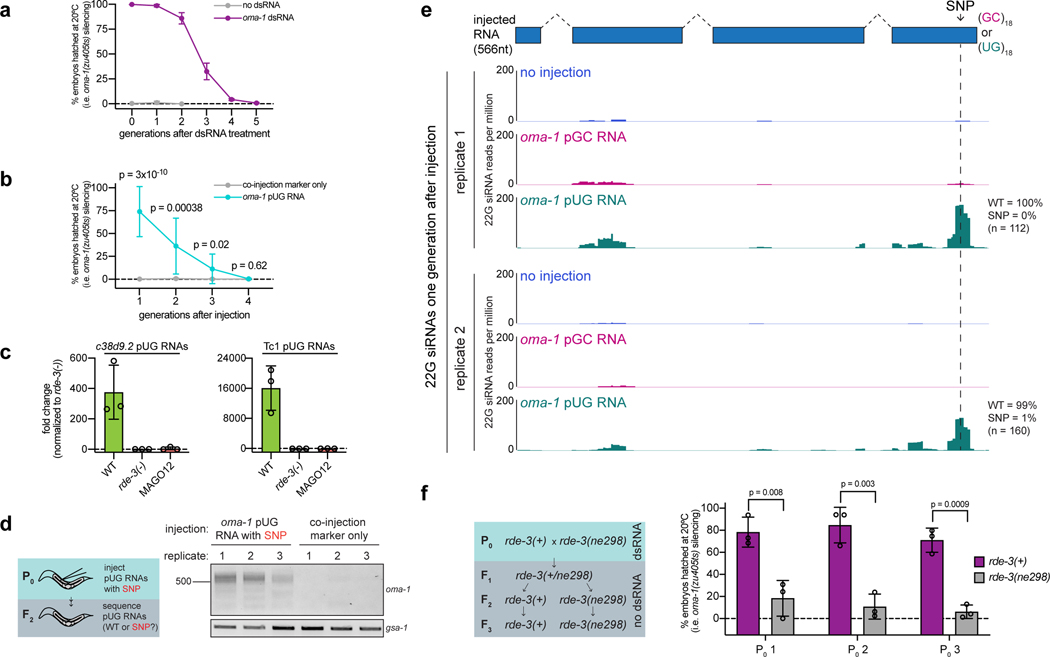
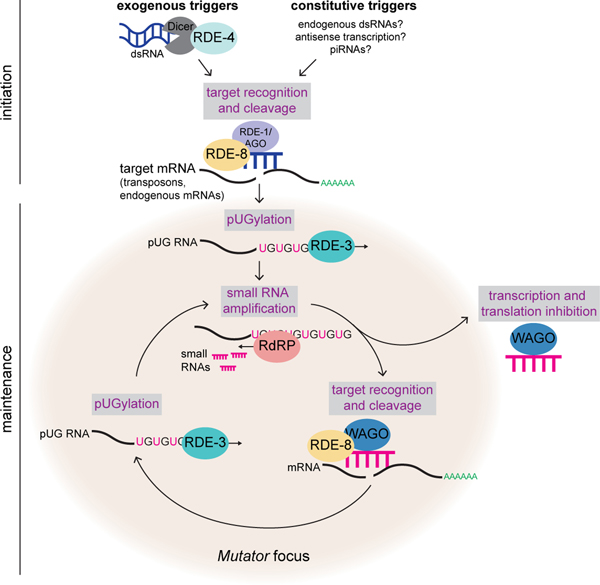
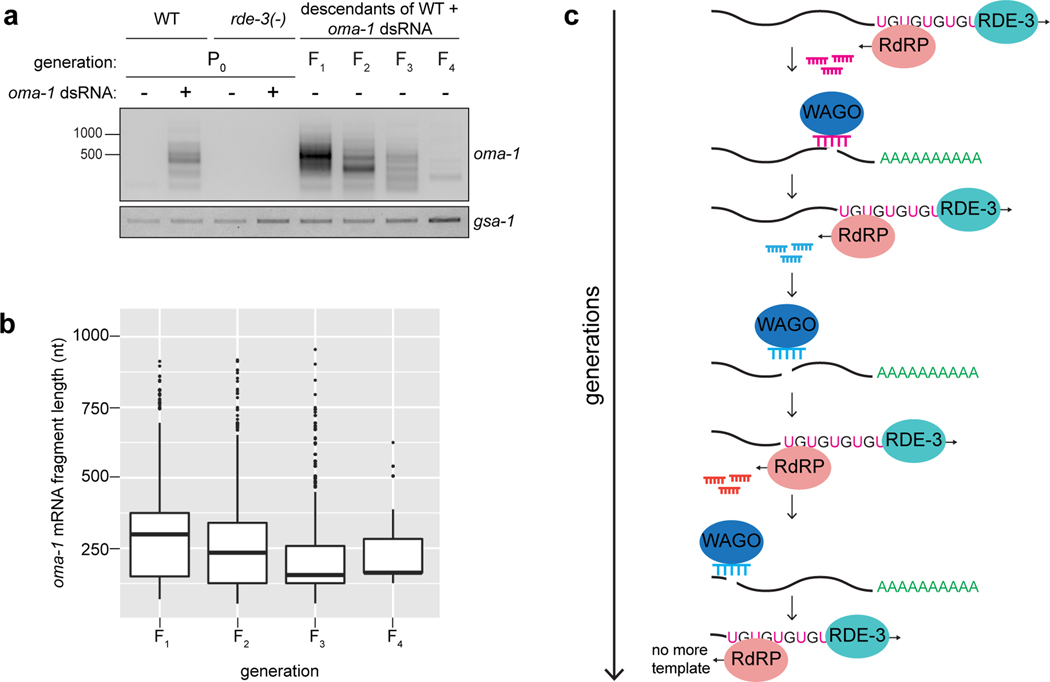
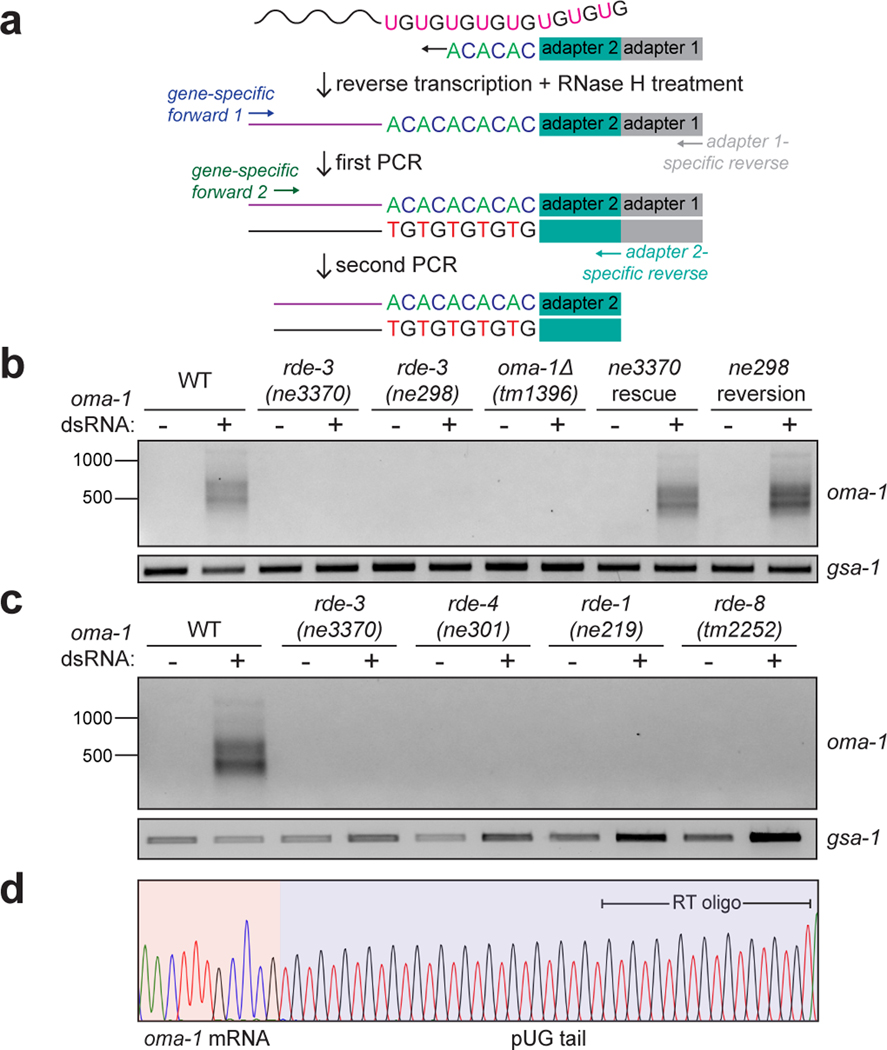
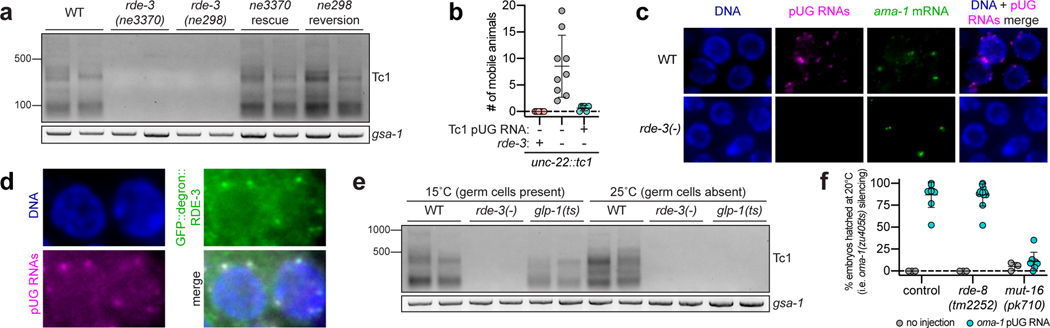
Mobile genetic elements threaten genome integrity in all organisms. RDE-3 (also known as MUT-2) is a ribonucleotidyltransferase that is required for transposon silencing and RNA interference in Caenorhabditis elegans1-4. When tethered to RNAs in heterologous expression systems, RDE-3 can add long stretches of alternating non-templated uridine (U) and guanosine (G) ribonucleotides to the 3' termini of these RNAs (designated poly(UG) or pUG tails)5. Here we show that, in its natural context in C. elegans, RDE-3 adds pUG tails to targets of RNA interference, as well as to transposon RNAs. RNA fragments attached to pUG tails with more than 16 perfectly alternating 3' U and G nucleotides become gene-silencing agents. pUG tails promote gene silencing by recruiting RNA-dependent RNA polymerases, which use pUG-tailed RNAs (pUG RNAs) as templates to synthesize small interfering RNAs (siRNAs). Our results show that cycles of pUG RNA-templated siRNA synthesis and siRNA-directed pUG RNA biogenesis underlie double-stranded-RNA-directed transgenerational epigenetic inheritance in the C. elegans germline. We speculate that this pUG RNA-siRNA silencing loop enables parents to inoculate progeny against the expression of unwanted or parasitic genetic elements.
Conflict of interest statement
Competing interests
M.W. has a patent (US20160145666A1) through Wisconsin Alumni Research Foundation (Madison, WI) for methods, kits, and compositions of matter relating to poly(UG) polymerases.
Figures


Comment in
-
Genes silenced down the generations, thanks to tails on messenger RNA.Nature. 2020 Jun;582(7811):191-192. doi: 10.1038/d41586-020-01417-2. Nature. 2020. PMID: 32433632 No abstract available.
References
Main references
-
- Collins J, Saari B. & Anderson P. Activation of a transposable element in the germ line but not the soma of Caenorhabditis elegans. Nature 328, 726–728 (1987). - PubMed
-
- Ketting RF, Haverkamp THA, van Luenen HGAM & Plasterk RHA mut-7 of C. elegans, Required for Transposon Silencing and RNA Interference, Is a Homolog of Werner Syndrome Helicase and RNaseD. Cell 99, 133–141 (1999). - PubMed
-
- Tabara H. et al. The rde-1 gene, RNA interference, and transposon silencing in C. elegans. Cell 99, 123–132 (1999). - PubMed
-
- Chen C-CG et al. A member of the polymerase beta nucleotidyltransferase superfamily is required for RNA interference in C. elegans. Curr. Biol 15, 378–383 (2005). - PubMed
Additional references
-
- Martin M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.journal 17, 10–12 (2011).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
