

Insights Into Genetic and Molecular Elements for Transgenic Crop Development
- PMID: 32499796
- PMCID: PMC7243915
- DOI: 10.3389/fpls.2020.00509
Insights Into Genetic and Molecular Elements for Transgenic Crop Development
Abstract
Climate change and the exploration of new areas of cultivation have impacted the yields of several economically important crops worldwide. Both conventional plant breeding based on planned crosses between parents with specific traits and genetic engineering to develop new biotechnological tools (NBTs) have allowed the development of elite cultivars with new features of agronomic interest. The use of these NBTs in the search for agricultural solutions has gained prominence in recent years due to their rapid generation of elite cultivars that meet the needs of crop producers, and the efficiency of these NBTs is closely related to the optimization or best use of their elements. Currently, several genetic engineering techniques are used in synthetic biotechnology to successfully improve desirable traits or remove undesirable traits in crops. However, the features, drawbacks, and advantages of each technique are still not well understood, and thus, these methods have not been fully exploited. Here, we provide a brief overview of the plant genetic engineering platforms that have been used for proof of concept and agronomic trait improvement, review the major elements and processes of synthetic biotechnology, and, finally, present the major NBTs used to improve agronomic traits in socioeconomically important crops.
Keywords: T-DNA delivery; minimal expression cassette; new biotechnological tools; plant genetic transformation; tissue culture.
Copyright © 2020 Basso, Arraes, Grossi-de-Sa, Moreira, Alves-Ferreira and Grossi-de-Sa.
Figures
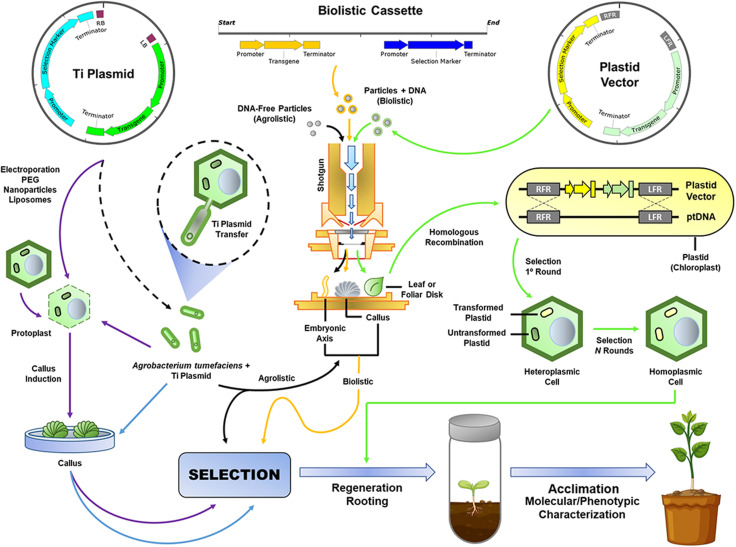
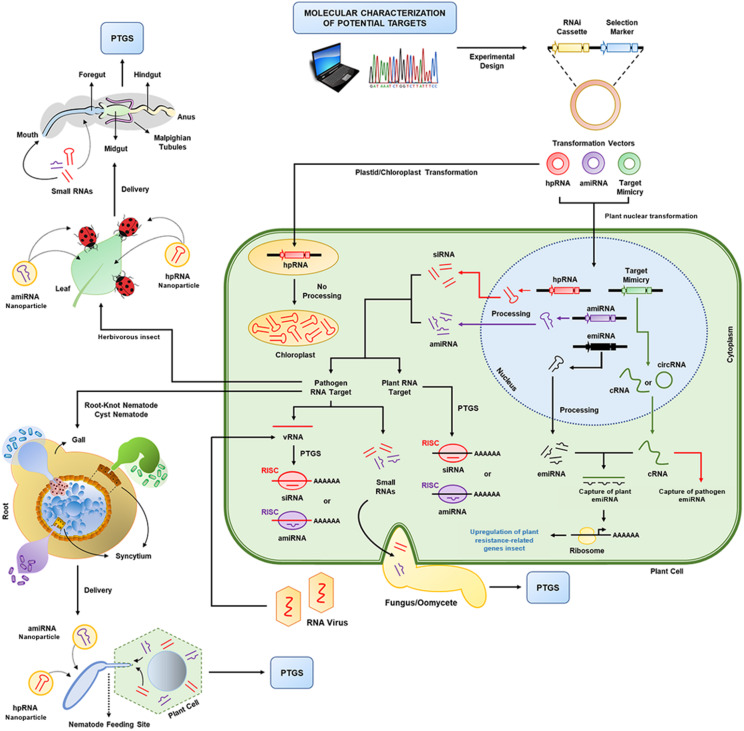
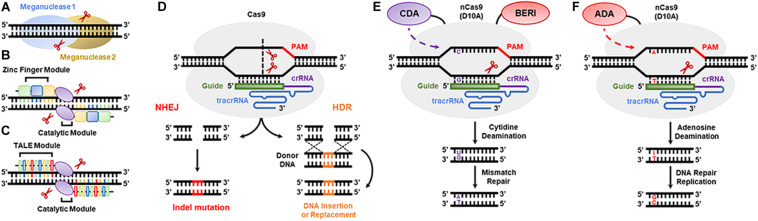
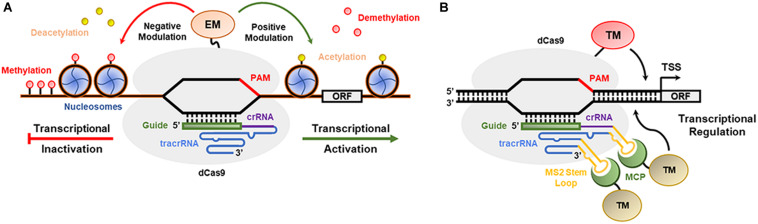
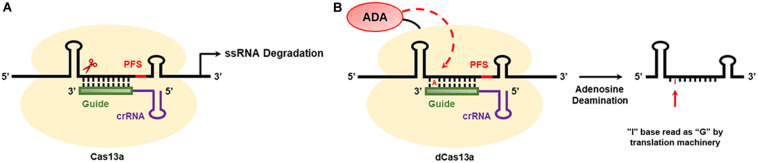
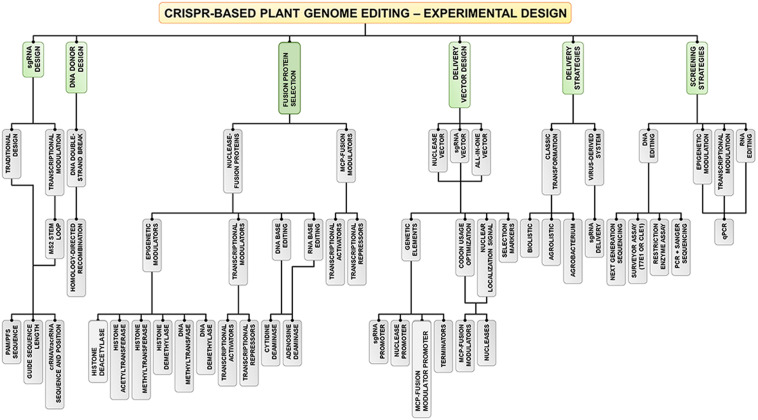
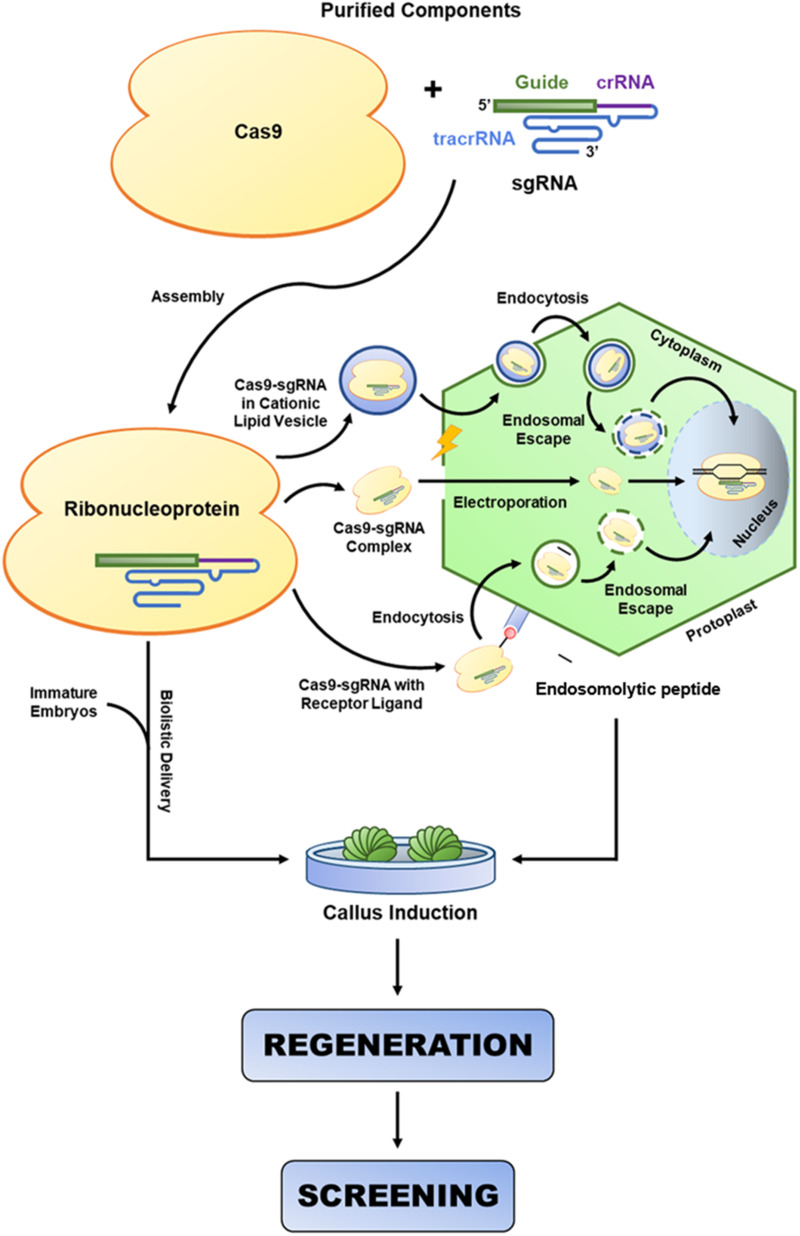
References
LinkOut - more resources
Full Text Sources
Other Literature Sources