

Highly active rubiscos discovered by systematic interrogation of natural sequence diversity
- PMID: 32500941
- PMCID: PMC7507306
- DOI: 10.15252/embj.2019104081
Highly active rubiscos discovered by systematic interrogation of natural sequence diversity
Abstract
CO2 is converted into biomass almost solely by the enzyme rubisco. The poor carboxylation properties of plant rubiscos have led to efforts that made it the most kinetically characterized enzyme, yet these studies focused on < 5% of its natural diversity. Here, we searched for fast-carboxylating variants by systematically mining genomic and metagenomic data. Approximately 33,000 unique rubisco sequences were identified and clustered into ≈ 1,000 similarity groups. We then synthesized, purified, and biochemically tested the carboxylation rates of 143 representatives, spanning all clusters of form-II and form-II/III rubiscos. Most variants (> 100) were active in vitro, with the fastest having a turnover number of 22 ± 1 s-1 -sixfold faster than the median plant rubisco and nearly twofold faster than the fastest measured rubisco to date. Unlike rubiscos from plants and cyanobacteria, the fastest variants discovered here are homodimers and exhibit a much simpler folding and activation kinetics. Our pipeline can be utilized to explore the kinetic space of other enzymes of interest, allowing us to get a better view of the biosynthetic potential of the biosphere.
Keywords: carbon fixation; carboxylation rate; enhanced photosynthesis; metagenomic survey; ribulose-1,5-bisphosphate carboxylase/oxygenase.
©2020 The Authors. Published under the terms of the CC BY 4.0 license.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

- A
A computational pipeline to extract rubisco sequences from genomic and metagenomic databases, cluster them based on sequence identity, and select representatives that cover the entire diversity of rubisco variants from nature.
- B
An experimental pipeline to screen the representative variants for carboxylation activity.
- C
Catalytic outliers are evaluated using radiometric assays for the determination of accurate k cat and K M carboxylation values as well as SC/O.


- A
Coupling rubisco activity to NADH oxidation is done by two enzymatic steps catalyzed by phosphoglucokinase (pgk) and glyceraldehyde 3‐phosphate dehydrogenase (gapdh) (Kubien et al, 2011). A gradient of CABP is used to gradually inhibit rubisco activity.
- B
NADH oxidation is monitored at 340 nm in a gas‐controlled plate reader under 4% CO2 and 0.2% O2 in order to favor carboxylation over oxygenation. The slope of the curves gives the rate of NADH oxidation, which is equal to twice the carboxylation rate; the rate with no CABP was measured in duplicates.
- C
Rate of carboxylation (y‐axis; slopes from panel B) as a function of CABP concentration (x‐axis). The x‐intercept gives the concentration of rubisco active sites ([E]) while the y‐intercept gives the reaction rate without CABP inhibition (V max); thus, the specific activity per active site is given by dividing V max by [E]; dashed line is a least‐square linear regression (r 2 > 0.99). For this example, rubisco from R. rubrum catalyzes ≈ 7 reactions per second.
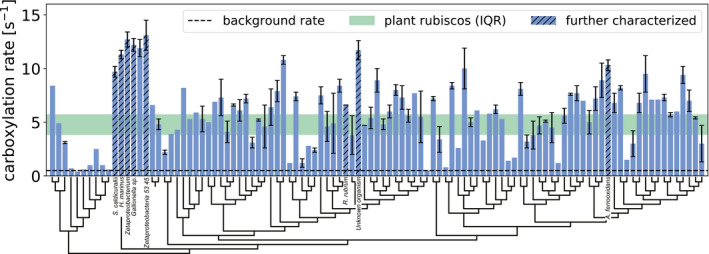

- A
Michaelis–Menten kinetic plots of rubiscos from Gallionella sp. (red; k cat = 22 ± 1.1 s−1; K M = 276 ± 6 μM) and S. elongatus (gray; k cat = 11.7 ± 0.6 s−1; K M = 200 ± 4 μM), measured by 14C labeling in the absence of oxygen at 25°C (see Materials and Methods); variants were measured in two biological repeats (circles and triangles), each in duplicate; dark and light green lines represent the median C3 (k cat = 3.1 s−1; K M = 14 μM) and C4 (k cat = 4.2 s−1; K M = 20 μM) plant rubiscos, respectively.
- B
k cat and K M values for all previously measured rubiscos (Flamholz et al, 2019) (gray) and the seven promising variants tested here (red for Gallionella sp. and blue for the other six); dashed line indicates the least‐square linear regression fit (slope in log scale is 0.4; r 2 = 0.44); histograms for k cat and K M are plotted on parallel axes and clearly show that the variants discovered in our screen are outliers (95th percentile) in both k cat and K M for CO2.
Comment in
-
Updated values for Table 1 of fastest rubisco carboxylation rates in Davidi et al 2020.EMBO J. 2025 May;44(9):2397-2399. doi: 10.1038/s44318-025-00419-y. Epub 2025 Apr 7. EMBO J. 2025. PMID: 40195502 Free PMC article.
References
-
- Aigner H, Wilson RH, Bracher A, Calisse L, Bhat JY, Hartl FU, Hayer‐Hartl M (2017) Plant RuBisCo assembly in E. coli with five chloroplast chaperones including BSD2. Science 358: 1272–1278 - PubMed
-
- Andralojc PJ, Madgwick PJ, Tao Y, Keys A, Ward JL, Beale MH, Loveland JE, Jackson PJ, Willis AC, Gutteridge S et al (2012) 2‐Carboxy‐D‐arabinitol 1‐phosphate (CA1P) phosphatase: evidence for a wider role in plant Rubisco regulation. Biochem J 442: 733–742 - PubMed
-
- Andrews TJ, Whitney SM (2003) Manipulating ribulose bisphosphate carboxylase/oxygenase in the chloroplasts of higher plants. Arch Biochem Biophys 414: 159–169 - PubMed
-
- Appia‐Ayme C, Quatrini R, Denis Y, Denizot F, Silver S, Roberto F, Veloso F, Valdés J, Pablo Cárdenas J, Esparza M et al (2006) Microarray and bioinformatic analyses suggest models for carbon metabolism in the autotroph Acidithiobacillus ferrooxidans . Hydrometallurgy 83: 273–280
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
