

Structural Dynamics and Topology of the Inactive Form of S21 Holin in a Lipid Bilayer Using Continuous-Wave Electron Paramagnetic Resonance Spectroscopy
- PMID: 32501696
- PMCID: PMC8011547
- DOI: 10.1021/acs.jpcb.0c03575
Structural Dynamics and Topology of the Inactive Form of S21 Holin in a Lipid Bilayer Using Continuous-Wave Electron Paramagnetic Resonance Spectroscopy
Abstract
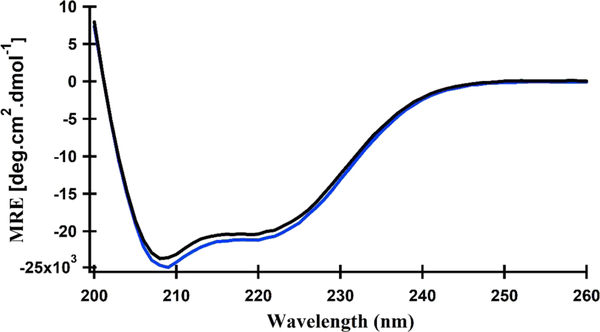
The bacteriophage infection cycle plays a crucial role in recycling the world's biomass. Bacteriophages devise various cell lysis systems to strictly control the length of the infection cycle for an efficient phage life cycle. Phages evolved with lysis protein systems, which can control and fine-tune the length of this infection cycle depending on the host and growing environment. Among these lysis proteins, holin controls the first and rate-limiting step of host cell lysis by permeabilizing the inner membrane at an allele-specific time and concentration hence known as the simplest molecular clock. Pinholin S21 is the holin from phage Φ21, which defines the cell lysis time through a predefined ratio of active pinholin and antipinholin (inactive form of pinholin). Active pinholin and antipinholin fine-tune the lysis timing through structural dynamics and conformational changes. Previously we reported the structural dynamics and topology of active pinholin S2168. Currently, there is no detailed structural study of the antipinholin using biophysical techniques. In this study, the structural dynamics and topology of antipinholin S2168IRS in DMPC proteoliposomes is investigated using electron paramagnetic resonance (EPR) spectroscopic techniques. Continuous-wave (CW) EPR line shape analysis experiments of 35 different R1 side chains of S2168IRS indicated restricted mobility of the transmembrane domains (TMDs), which were predicted to be inside the lipid bilayer when compared to the N- and C-termini R1 side chains. In addition, the R1 accessibility test performed on 24 residues using the CW-EPR power saturation experiment indicated that TMD1 and TMD2 of S2168IRS were incorporated into the lipid bilayer where N- and C-termini were located outside of the lipid bilayer. Based on this study, a tentative model of S2168IRS is proposed where both TMDs remain incorporated into the lipid bilayer and N- and C-termini are located outside of the lipid bilayer. This work will pave the way for the further studies of other holins using biophysical techniques and will give structural insights into these biological clocks in molecular detail.
Conflict of interest statement
The authors declare no competing financial interest.
Figures








References
-
- Young R Bacteriophage holins: Deadly diversity. J. Mol. Microbiol. Biotechnol 2002, 4, 21–36. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources