

Oral epithelial IL-22/STAT3 signaling licenses IL-17-mediated immunity to oral mucosal candidiasis
- PMID: 32503875
- PMCID: PMC7340112
- DOI: 10.1126/sciimmunol.aba0570
Oral epithelial IL-22/STAT3 signaling licenses IL-17-mediated immunity to oral mucosal candidiasis
Abstract
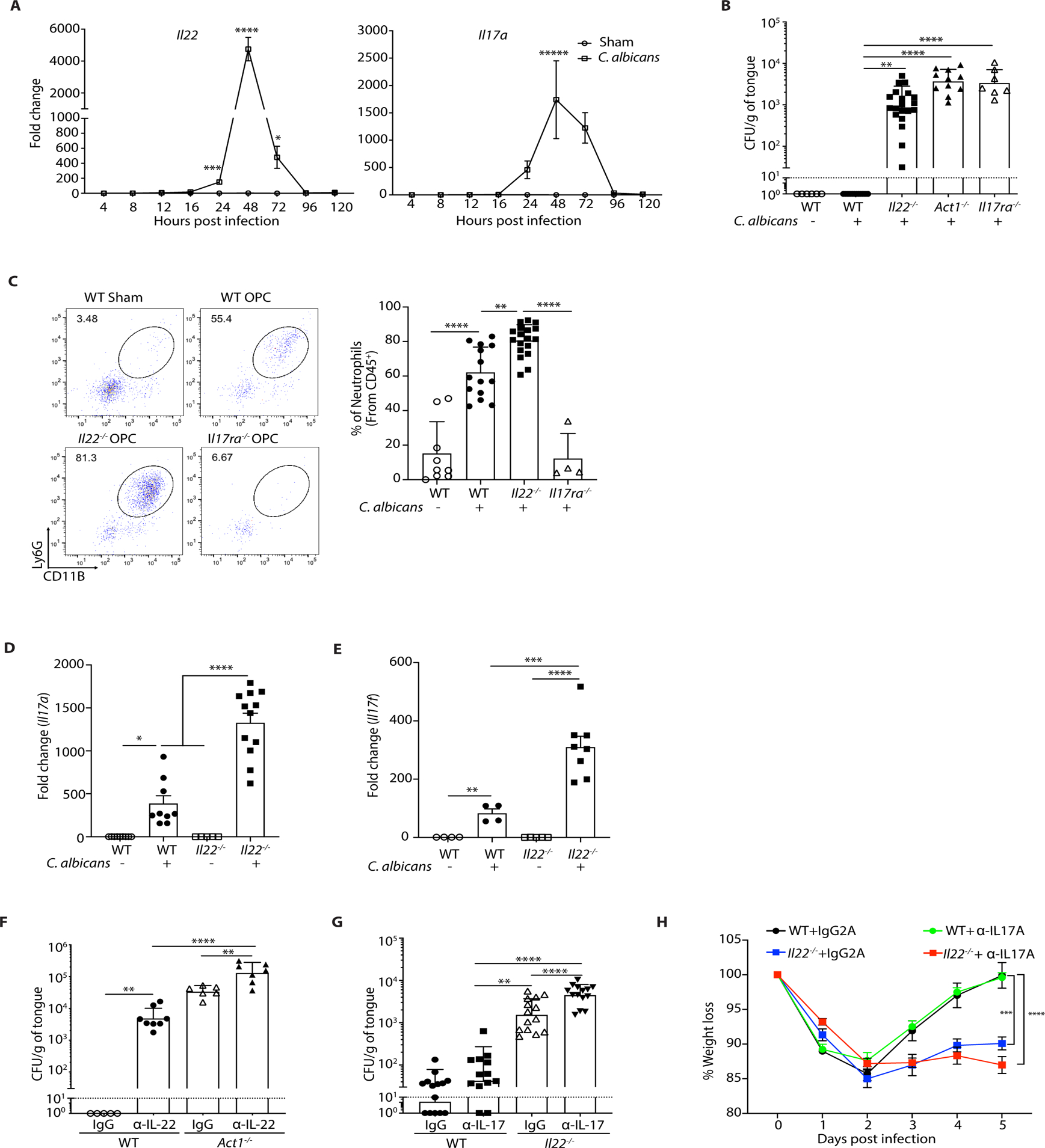
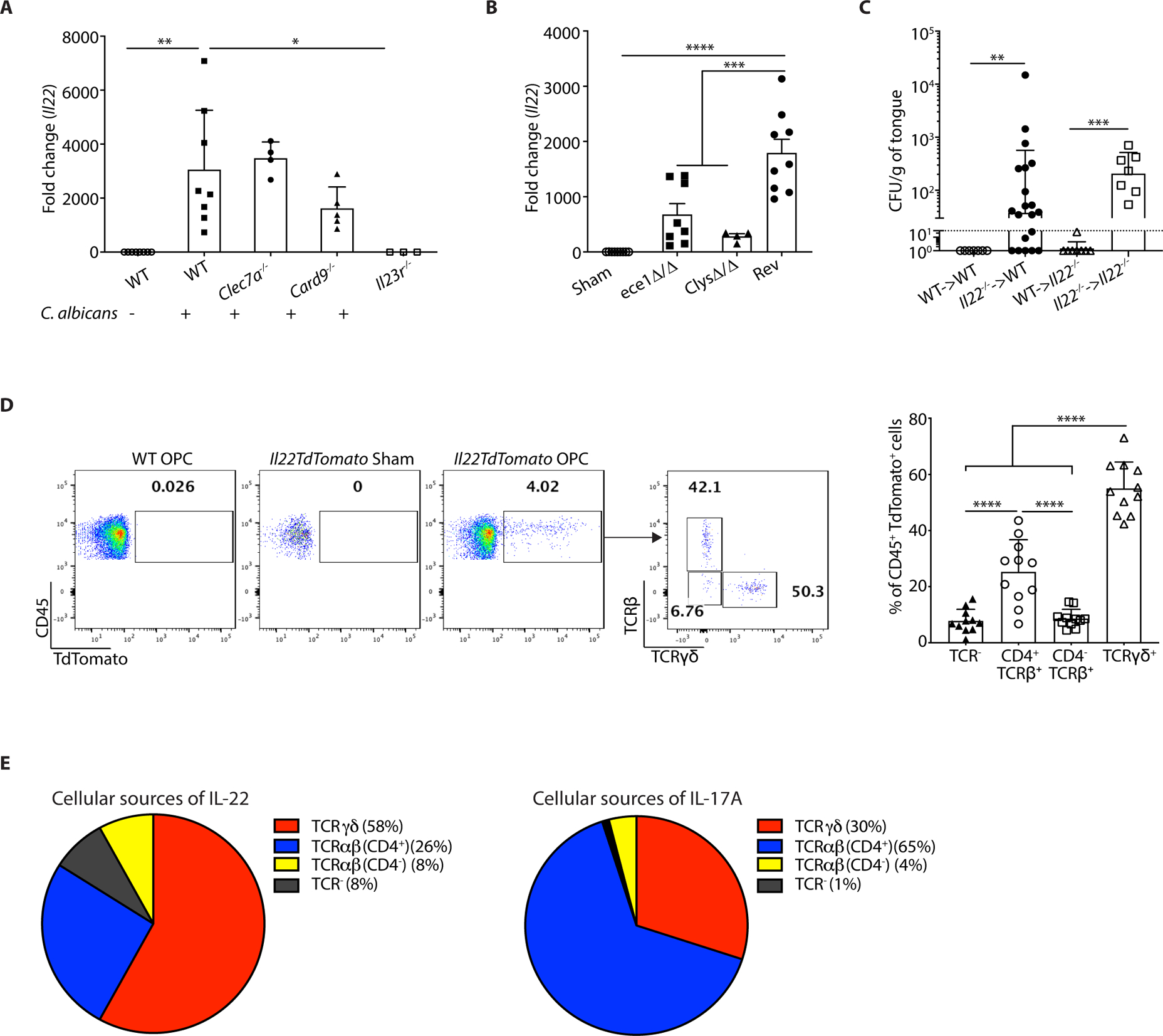
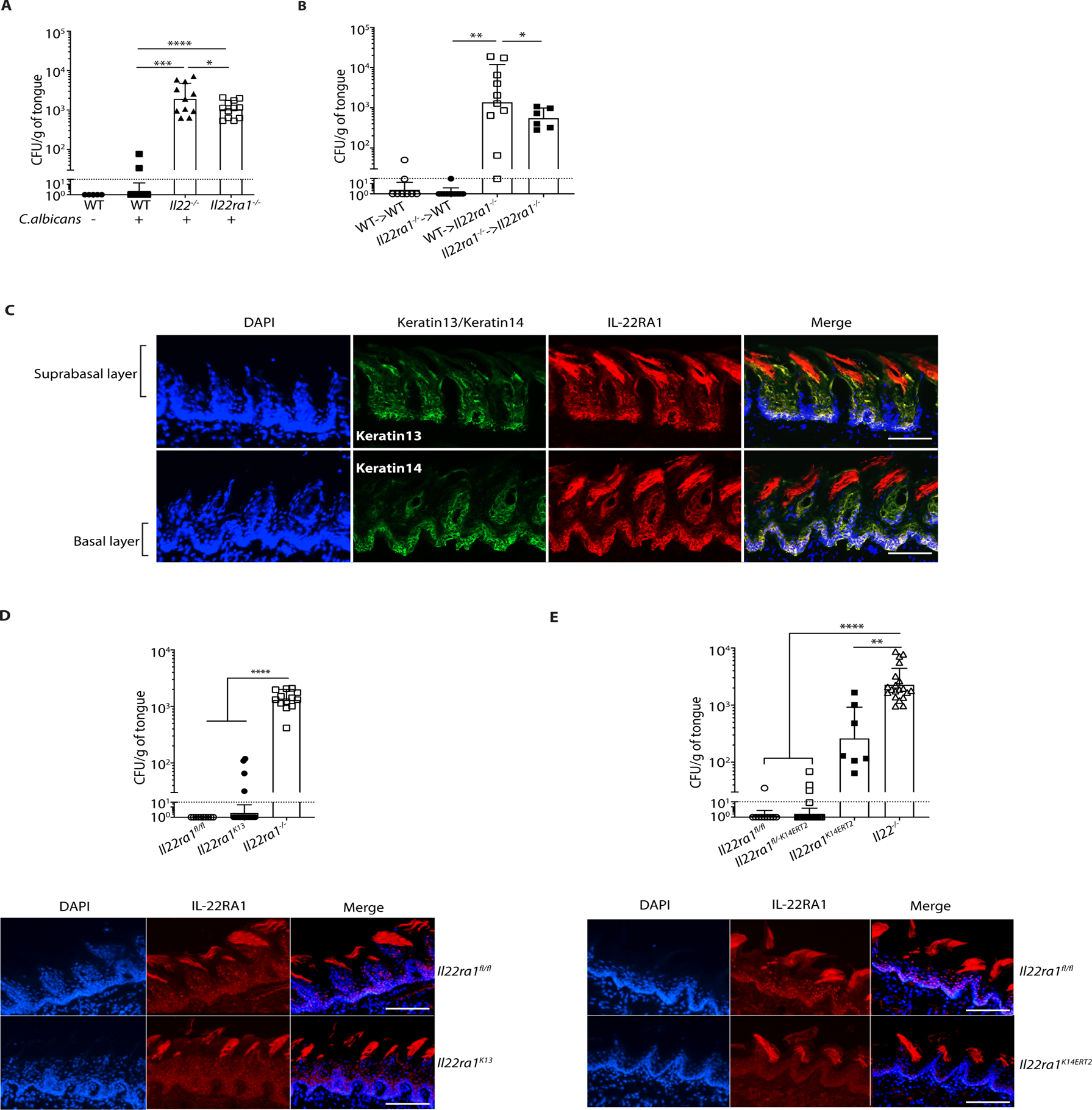
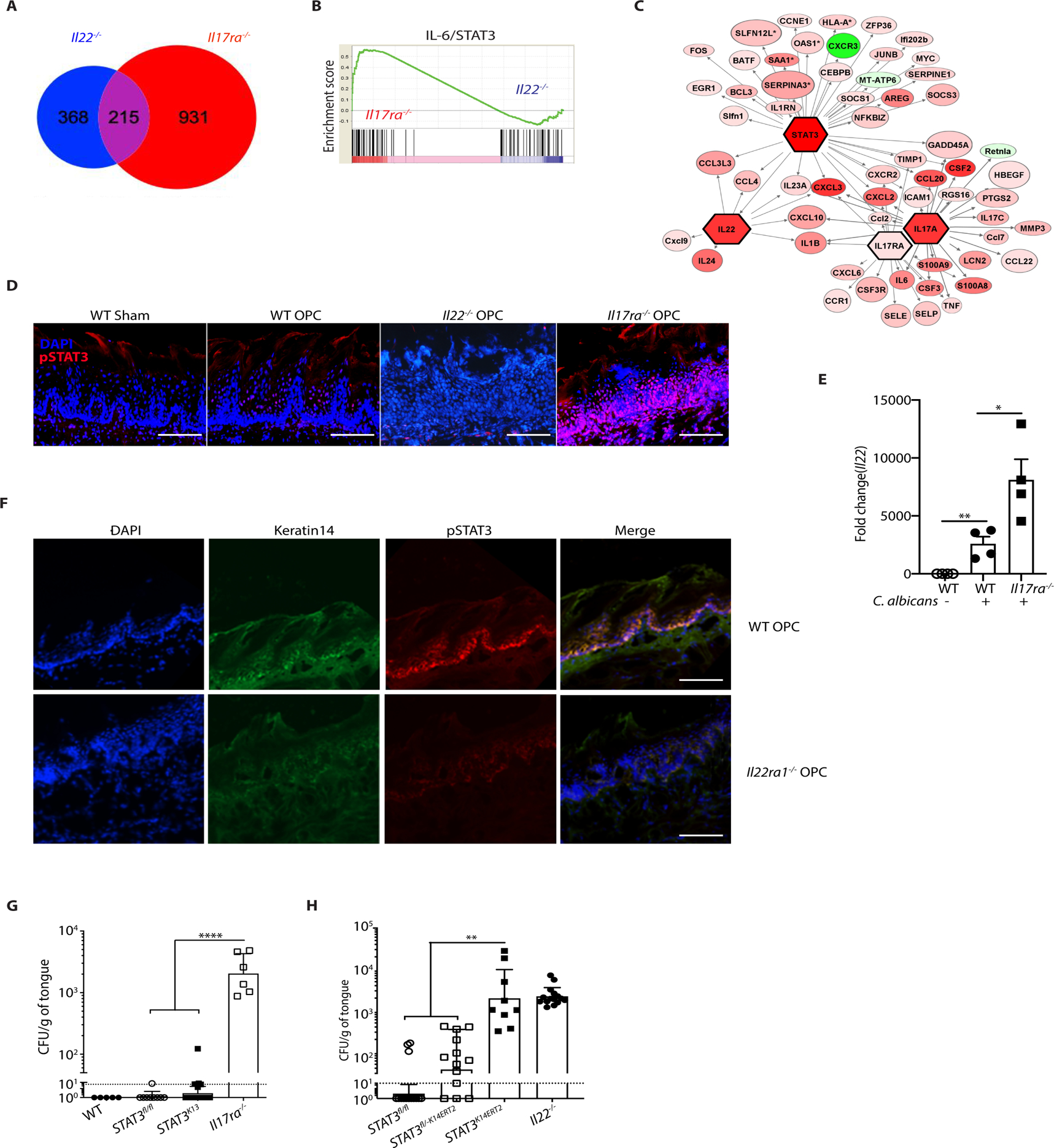
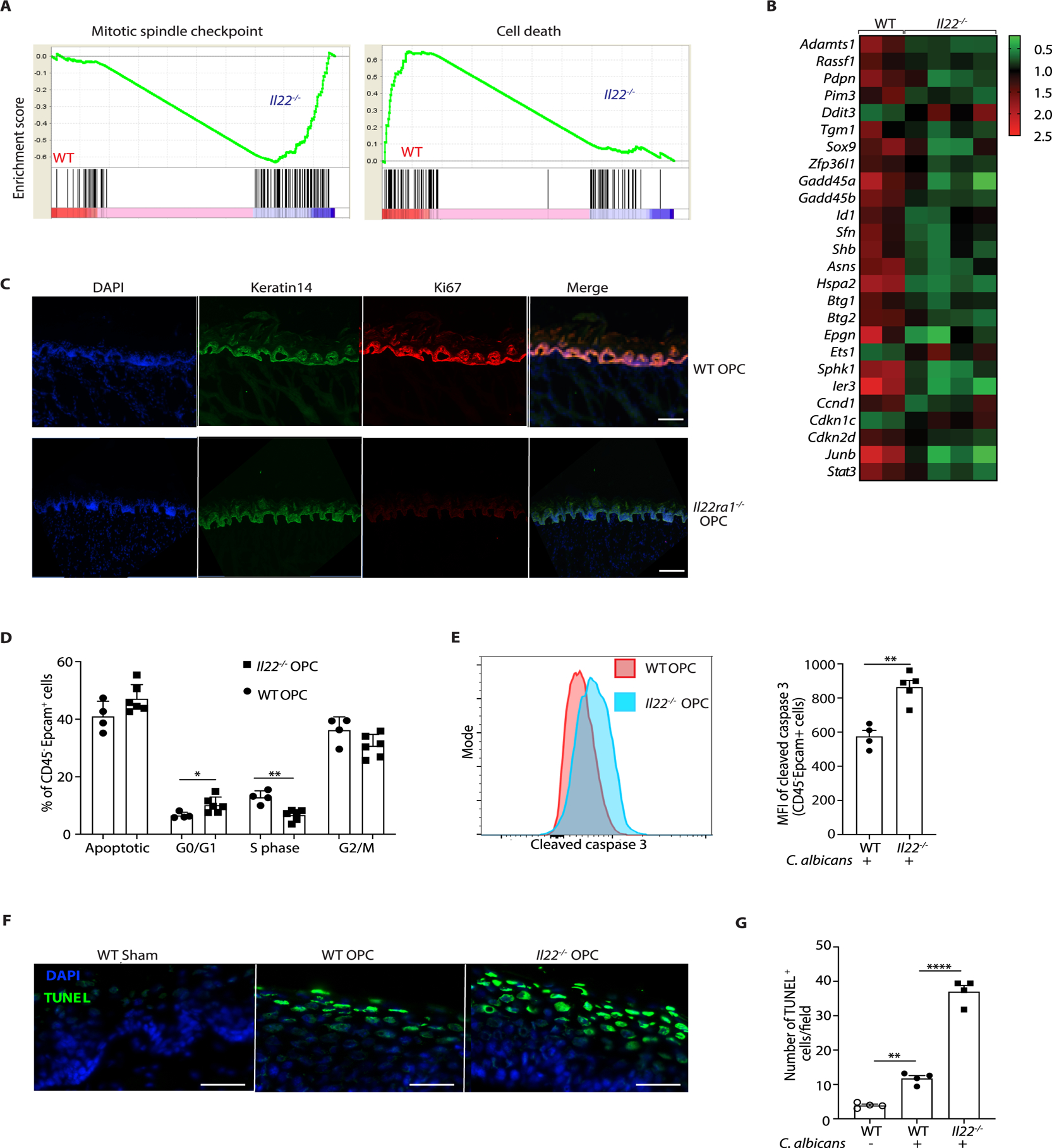
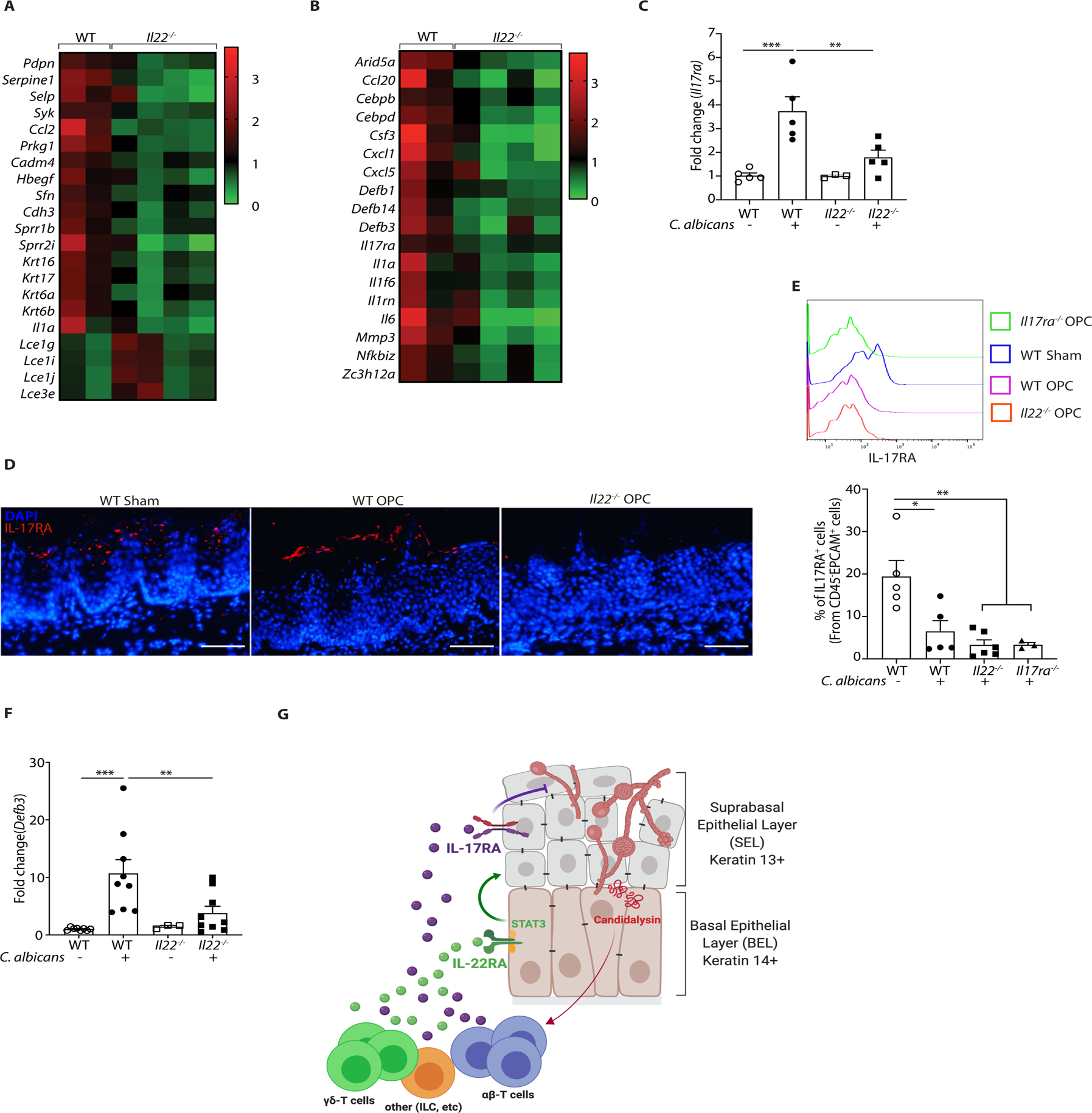
Oropharyngeal candidiasis (OPC; thrush) is an opportunistic infection caused by the commensal fungus Candida albicans Interleukin-17 (IL-17) and IL-22 are cytokines produced by type 17 lymphocytes. Both cytokines mediate antifungal immunity yet activate quite distinct downstream signaling pathways. While much is now understood about how IL-17 promotes immunity in OPC, the activities of IL-22 are far less well delineated. We show that, despite having similar requirements for induction from type 17 cells, IL-22 and IL-17 function nonredundantly during OPC. We find that the IL-22 and IL-17 receptors are required in anatomically distinct locations within the oral mucosa; loss of IL-22RA1 or signal transducer and activator of transcription 3 (STAT3) in the oral basal epithelial layer (BEL) causes susceptibility to OPC, whereas IL-17RA is needed in the suprabasal epithelial layer (SEL). Transcriptional profiling of the tongue linked IL-22/STAT3 not only to oral epithelial cell proliferation and survival but also, unexpectedly, to driving an IL-17-specific gene signature. We show that IL-22 mediates regenerative signals on the BEL that replenish the IL-17RA-expressing SEL, thereby restoring the ability of the oral epithelium to respond to IL-17 and thus to mediate antifungal events. Consequently, IL-22 signaling in BEL "licenses" IL-17 signaling in the oral mucosa, revealing spatially distinct yet cooperative activities of IL-22 and IL-17 in oral candidiasis.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
COMPETING INTERESTS
SLG previously received a grant from Janssen to evaluate anti-IL-17A antibody function in OPC (110). Candidalysin has been patented by King’s College London, UK and Hans-Knöll Institut, Jena, DE (US Patent No.: 9,969,796; EU Patent No.: 2984103). The other authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- R35 HL139930/HL/NHLBI NIH HHS/United States
- R01 DE022550/DE/NIDCR NIH HHS/United States
- R37 DE022550/DE/NIDCR NIH HHS/United States
- R01 DK104680/DK/NIDDK NIH HHS/United States
- R01 AI107825/AI/NIAID NIH HHS/United States
- S10 OD011925/OD/NIH HHS/United States
- R01 AR060744/AR/NIAMS NIH HHS/United States
- U19 AI110820/AI/NIAID NIH HHS/United States
- R01 DE023815/DE/NIDCR NIH HHS/United States
- K01 HL135476/HL/NHLBI NIH HHS/United States
- R03 HL154231/HL/NHLBI NIH HHS/United States
- WT_/Wellcome Trust/United Kingdom
- R01 AR071720/AR/NIAMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
