

Prefoldin-like Bud27 influences the transcription of ribosomal components and ribosome biogenesis in Saccharomyces cerevisiae
- PMID: 32503921
- PMCID: PMC7491330
- DOI: 10.1261/rna.075507.120
Prefoldin-like Bud27 influences the transcription of ribosomal components and ribosome biogenesis in Saccharomyces cerevisiae
Abstract
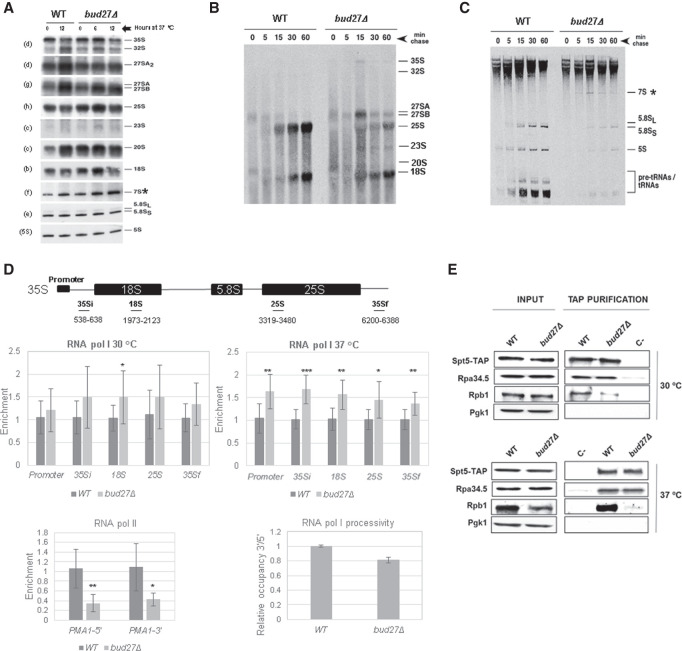
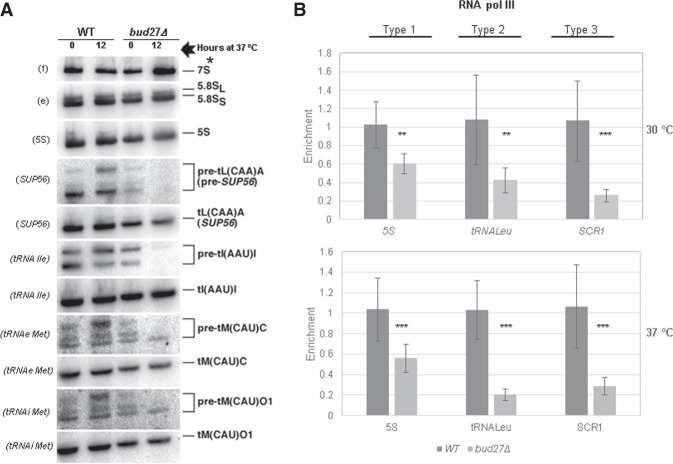
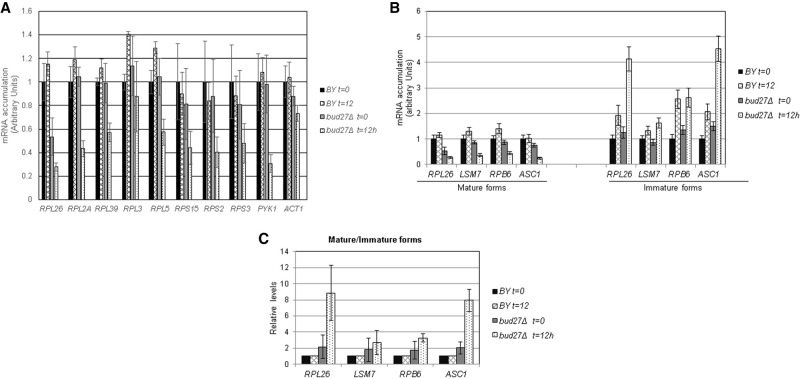
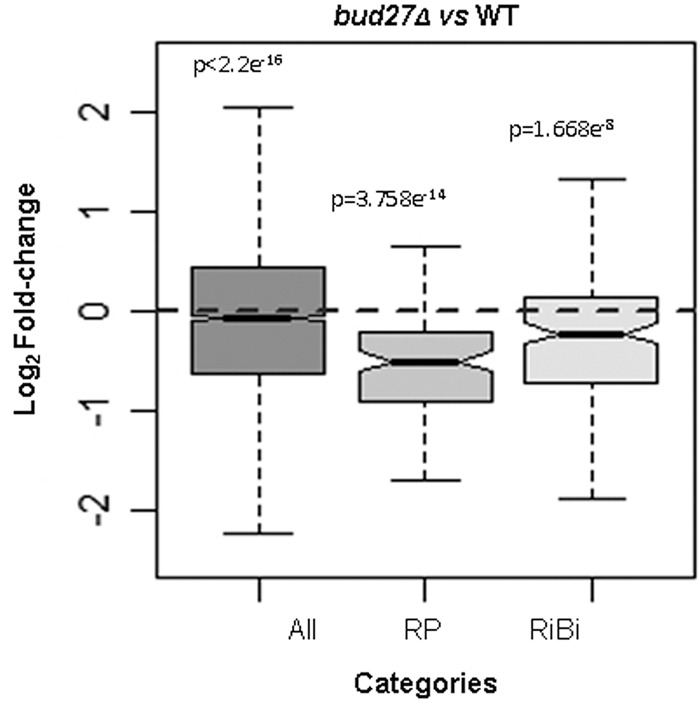
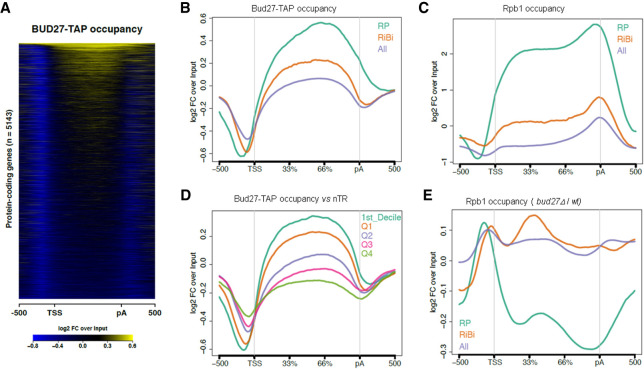
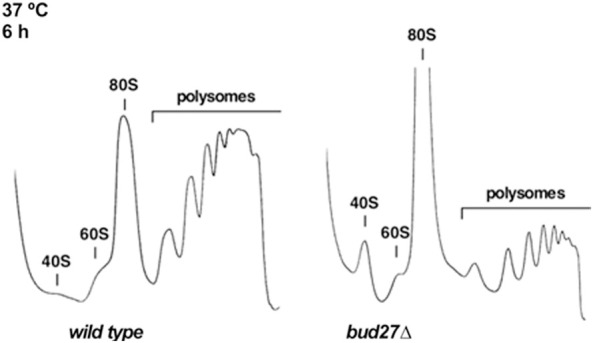
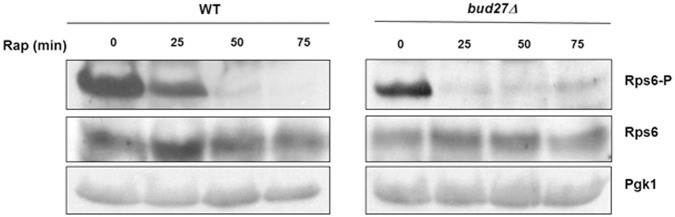
Understanding the functional connection that occurs for the three nuclear RNA polymerases to synthesize ribosome components during the ribosome biogenesis process has been the focal point of extensive research. To preserve correct homeostasis on the production of ribosomal components, cells might require the existence of proteins that target a common subunit of these RNA polymerases to impact their respective activities. This work describes how the yeast prefoldin-like Bud27 protein, which physically interacts with the Rpb5 common subunit of the three RNA polymerases, is able to modulate the transcription mediated by the RNA polymerase I, likely by influencing transcription elongation, the transcription of the RNA polymerase III, and the processing of ribosomal RNA. Bud27 also regulates both RNA polymerase II-dependent transcription of ribosomal proteins and ribosome biogenesis regulon genes, likely by occupying their DNA ORFs, and the processing of the corresponding mRNAs. With RNA polymerase II, this association occurs in a transcription rate-dependent manner. Our data also indicate that Bud27 inactivation alters the phosphorylation kinetics of ribosomal protein S6, a readout of TORC1 activity. We conclude that Bud27 impacts the homeostasis of the ribosome biogenesis process by regulating the activity of the three RNA polymerases and, in this way, the synthesis of ribosomal components. This quite likely occurs through a functional connection of Bud27 with the TOR signaling pathway.
Keywords: RNA polymerases; Saccharomyces cerevisiae; prefoldin-like; ribosome biogenesis; transcription.
© 2020 Martínez-Fernández et al.; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures

Similar articles
-
A High-Copy Suppressor Screen Reveals a Broad Role of Prefoldin-like Bud27 in the TOR Signaling Pathway in Saccharomyces cerevisiae.Genes (Basel). 2022 Apr 24;13(5):748. doi: 10.3390/genes13050748. Genes (Basel). 2022. PMID: 35627133 Free PMC article.
-
The prefoldin bud27 mediates the assembly of the eukaryotic RNA polymerases in an rpb5-dependent manner.PLoS Genet. 2013;9(2):e1003297. doi: 10.1371/journal.pgen.1003297. Epub 2013 Feb 14. PLoS Genet. 2013. PMID: 23459708 Free PMC article.
-
The Yeast Prefoldin Bud27.Adv Exp Med Biol. 2018;1106:109-118. doi: 10.1007/978-3-030-00737-9_8. Adv Exp Med Biol. 2018. PMID: 30484156 Review.
-
Yeast Bud27 modulates the biogenesis of Rpc128 and Rpc160 subunits and the assembly of RNA polymerase III.Biochim Biophys Acta. 2015 Nov;1849(11):1340-53. doi: 10.1016/j.bbagrm.2015.09.010. Epub 2015 Sep 28. Biochim Biophys Acta. 2015. PMID: 26423792
-
Feedback regulation of ribosome assembly.Curr Genet. 2018 Apr;64(2):393-404. doi: 10.1007/s00294-017-0764-x. Epub 2017 Oct 11. Curr Genet. 2018. PMID: 29022131 Review.
Cited by
-
A High-Copy Suppressor Screen Reveals a Broad Role of Prefoldin-like Bud27 in the TOR Signaling Pathway in Saccharomyces cerevisiae.Genes (Basel). 2022 Apr 24;13(5):748. doi: 10.3390/genes13050748. Genes (Basel). 2022. PMID: 35627133 Free PMC article.
-
Regulation of Eukaryotic RNAPs Activities by Phosphorylation.Front Mol Biosci. 2021 Jun 25;8:681865. doi: 10.3389/fmolb.2021.681865. eCollection 2021. Front Mol Biosci. 2021. PMID: 34250017 Free PMC article. Review.
-
RNA Pol II Assembly Affects ncRNA Expression.Int J Mol Sci. 2023 Dec 29;25(1):507. doi: 10.3390/ijms25010507. Int J Mol Sci. 2023. PMID: 38203678 Free PMC article.
-
Prefoldin Subunits and Its Associate Partners: Conservations and Specificities in Plants.Plants (Basel). 2024 Feb 18;13(4):556. doi: 10.3390/plants13040556. Plants (Basel). 2024. PMID: 38498526 Free PMC article. Review.
-
Transcription by the Three RNA Polymerases under the Control of the TOR Signaling Pathway in Saccharomyces cerevisiae.Biomolecules. 2023 Apr 3;13(4):642. doi: 10.3390/biom13040642. Biomolecules. 2023. PMID: 37189389 Free PMC article. Review.
References
-
- Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K. 1994. Current protocols in molecular biology, pp. 13.10.11–13.14.17. Wiley, New York.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases